Птицы — класс теплокровных яйцекладущих животных, приспособившихся к полету. Самый многочисленный по числу видов, включает около 11 тысяч видов.

Их форма тела, легкость, перья — весь организм птиц создан для полета! Вы убедитесь в этом, изучая строении пищеварительной, дыхательной, нервной и других систем. Птицы расселились по всему миру, и смогли благодаря теплокровности (в отличие от амфибий и рептилий)
освоить зоны с холодным климатом.
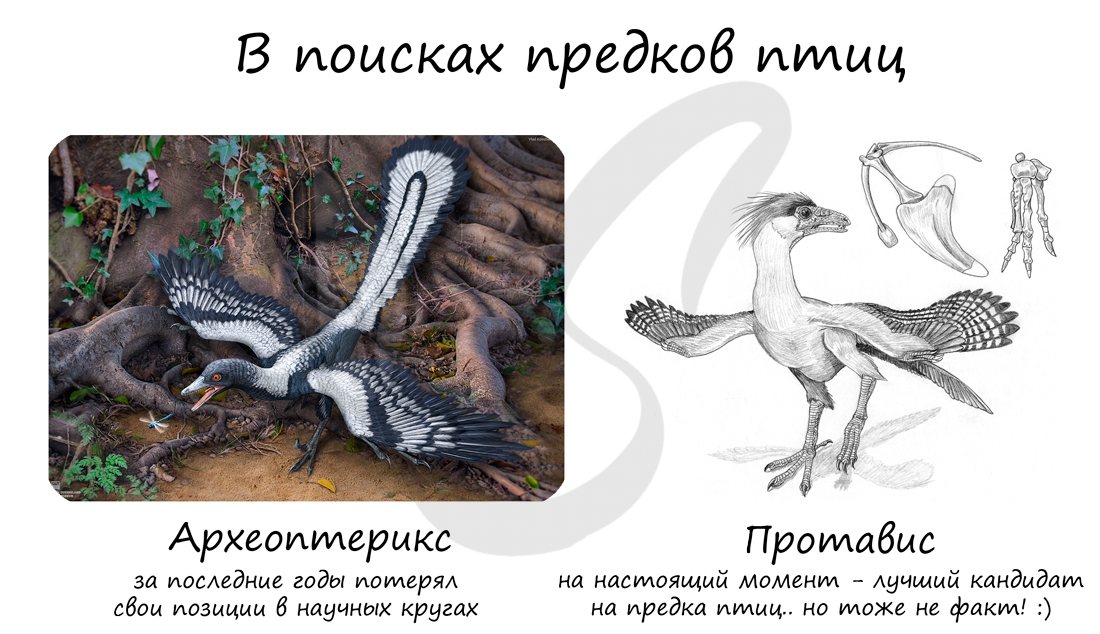
Птицы произошли от древних летающих пресмыкающихся — археоптериксов. Об этом свидетельствует ряд общих признаков: строение
яиц, наличие грудной клетки, сухая кожа без желез, перья птиц являются производными роговых чешуй (которые покрывали тело
рептилий).
Преимущества птицам дали появившиеся у них прогрессивные черты строения — ароморфозы:
Ароморфозы птиц
- Перьевой покров
- Полное разделение кругов кровообращения и четырехкамерное сердце
- Двойное дыхание и губчатые легкие
Перья птиц представляют собой видоизмененную чешую рептилий. Тело птиц покрыто перьями — роговыми наружными образованиями,
состоящими из полого стержня, нижняя часть которого называется очин,
с пушистыми плотно прилегающими друг к другу отростками по бокам (опахало).
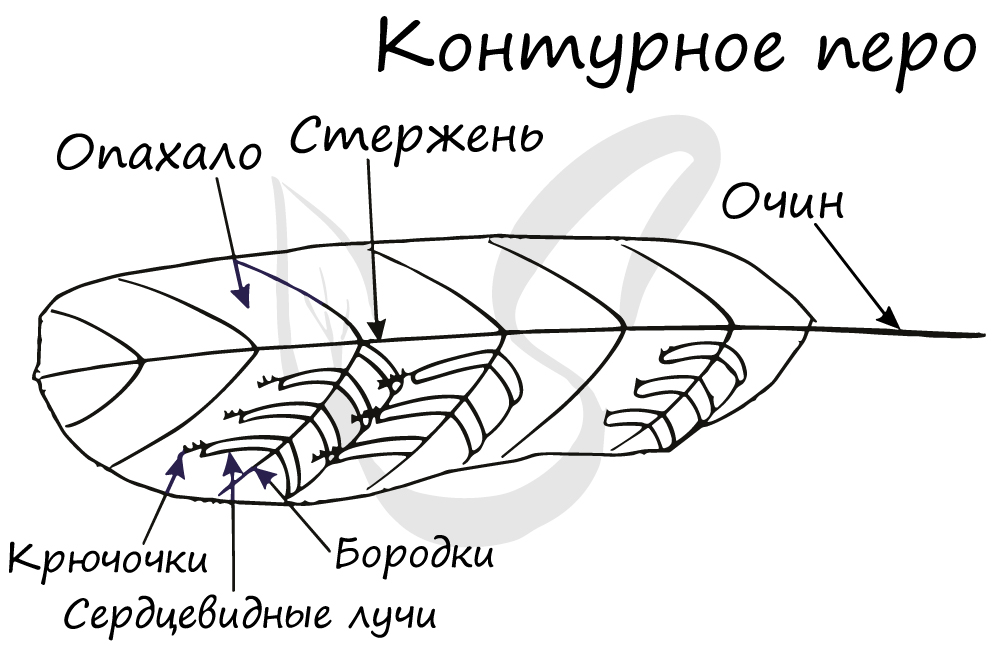
Опахало состоит из бородок 1-ого порядка, которые крепятся напрямую к стержню, и бородок 2-ого порядка, расположенных на
бородках 1-ого порядка.
Очин погружен в кожу, а стержень с опахалом расположены снаружи над поверхностью тела. Перьевой покров придает птице
характерную обтекаемую форму тела. Надо заметить, что контурные — не единственные перья птиц.
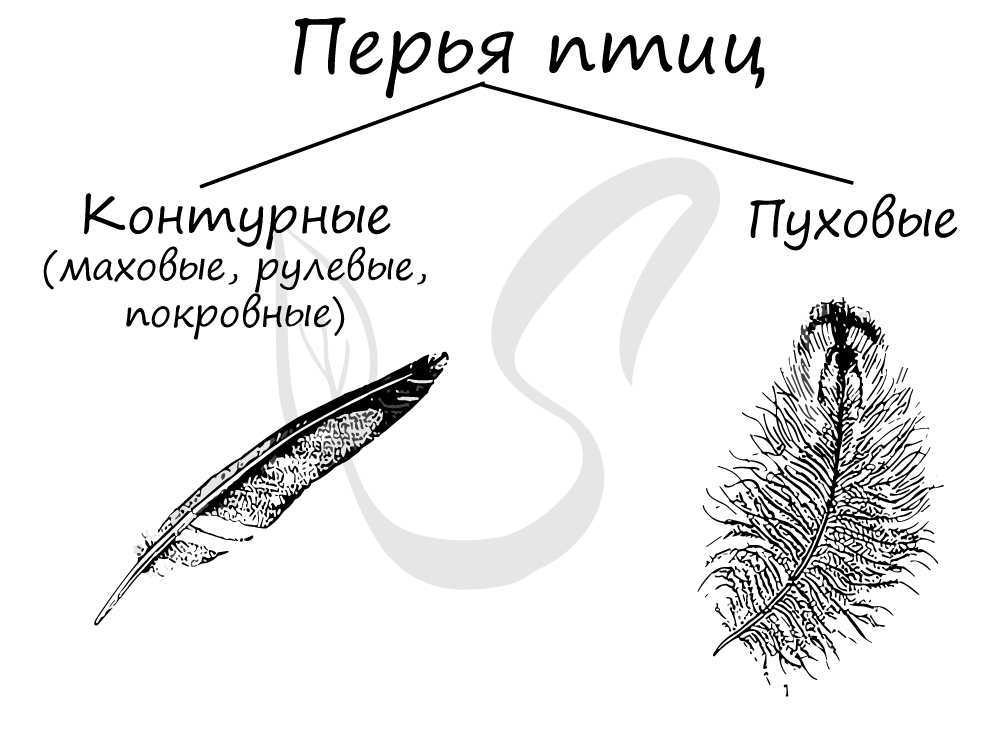
Маховые перья растут по краю крыла, именно они поддерживают птицу во время полета. Рулевые перья являются продолжением хвоста у большинства
птиц, играют важную роль при изменении направления полета. Покровные перья формируют характерный обтекаемый контур птицы, за счет чего
и получили свое название.
По-иному устроены пуховые перья: их бородки не несут крючочков, вследствие чего плотно сцепленное опахало не образуется.
Пуховые перья равномерно покрывают все тело птицы, обеспечивая теплоизоляцию от окружающей среды. Периодически у птиц происходит линька —
смена оперения.
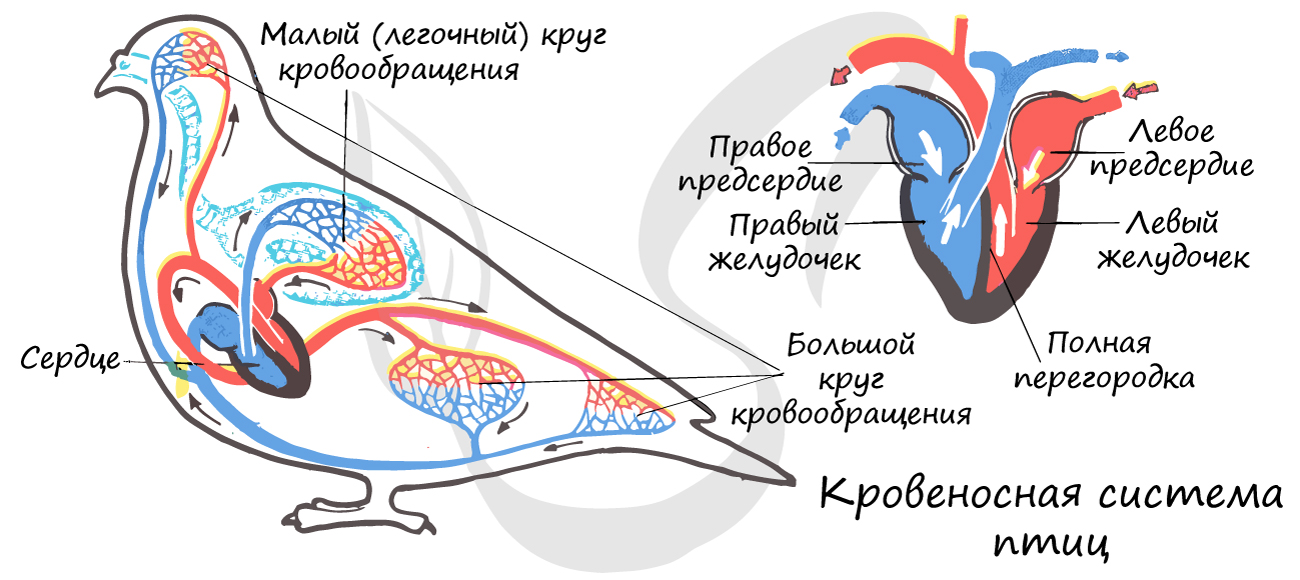
У птиц имеется полная перегородка в сердце, разделяющая его камеру на два изолированных желудочка. Кровь больше не смешивается:
два круга кровообращения полностью отделены друг от друга.
Уровень обмена веществ становится гораздо выше, чем у земноводных и пресмыкающихся, что проявляется в приобретении птицами теплокровности (гомойотермии). Температура тела птиц более не зависит от окружающей среды, они могут осваивать среды обитания с низкими температурами.

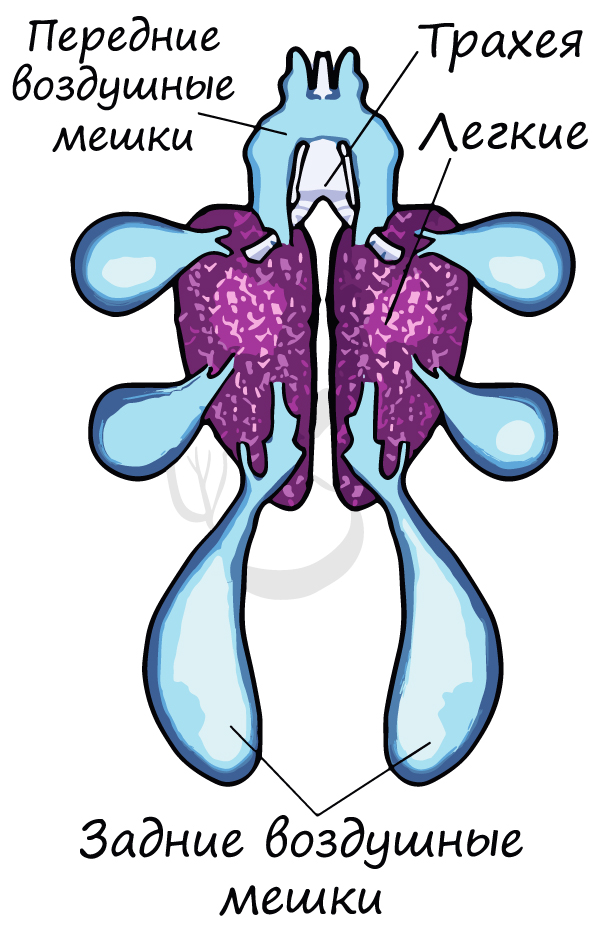
За счет развития легочных мешков — воздухоносных полостей, связанных с дыхательной системой, у птиц газообмен в легких
осуществляется и на вдохе, и на выдохе.

Для птиц характерны более сложноустроенные губчатые легкие, дыхательная площадь которых превышает площадь ячеистых легких
пресмыкающихся. Газообмен в них происходит с большей эффективностью.
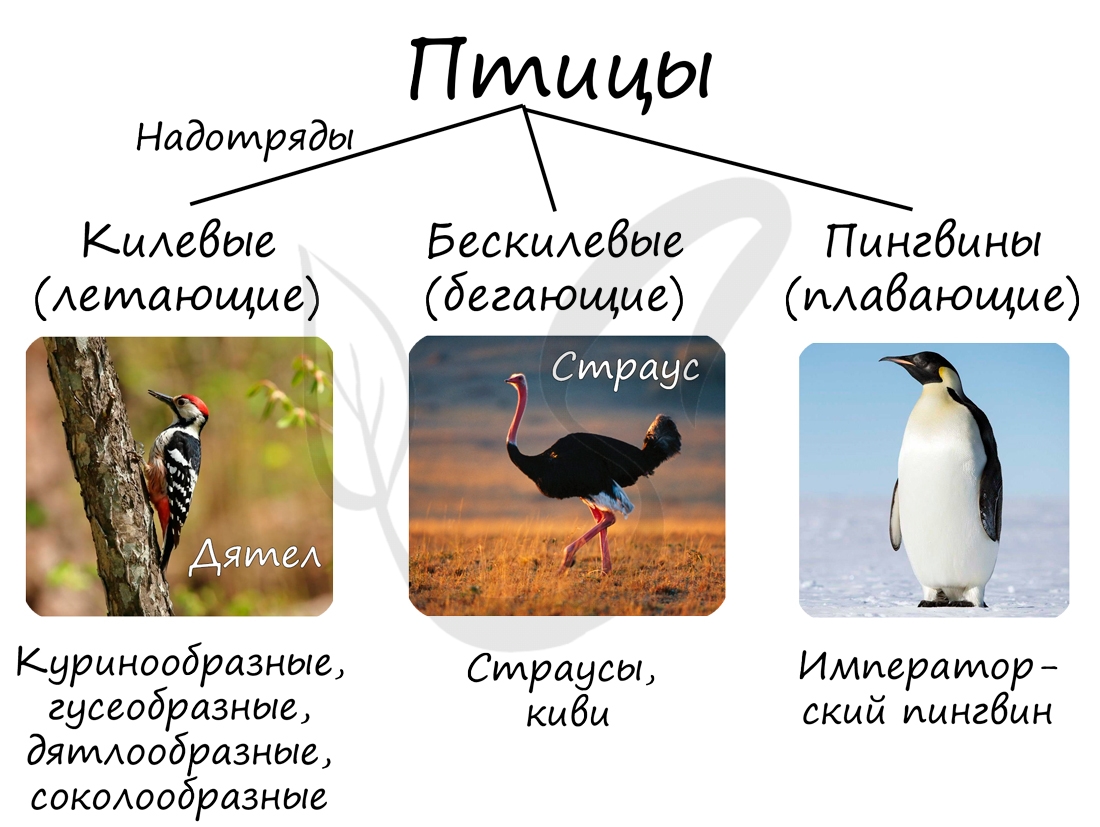
Чтобы основательно изучить тот или иной раздел зоологии очень важно знать классификацию. Без классификаций в голове будет сплошная
«каша», так что относитесь к ним с особым вниманием. Класс птицы состоит из трех надотрядов: бескилевые, килевые и пингвины.
Мы начнем подробное знакомство с птицами, взяв за основу известного нам типичного представителя — сизого голубя, относящегося
к надотряду килевые.
Сизый голубь
- Покровы, опорно-двигательная система
- Пищеварительная система
- Дыхательная система
- Кровеносная система
- Выделительная система
- Нервная система
- Половая система
Тело покрыто перьями, имеет обтекаемую, аэродинамическую форму. Оно подразделяется на голову, шею, туловище и хвост.
Передние конечности в виде крыльев, задние — ноги с 4 пальцами.
На голове хорошо заметен клюв, включающий надклювье и подклювье. У хищных птиц клюв приобретает заостренную форму,
для того чтобы отрывать им плоть.

Сухая кожа практически полностью лишена желез. Единственная железа, копчиковая, расположена над хвостовыми позвонками. Секрет
копчиковой железы имеет маслянистый характер, используется птицей для смазки крыльев, предотвращая их намокание. Особенно
хорошо развита эта железа у водоплавающих птиц: они смазывают ее секретом плотный перьевой покров.

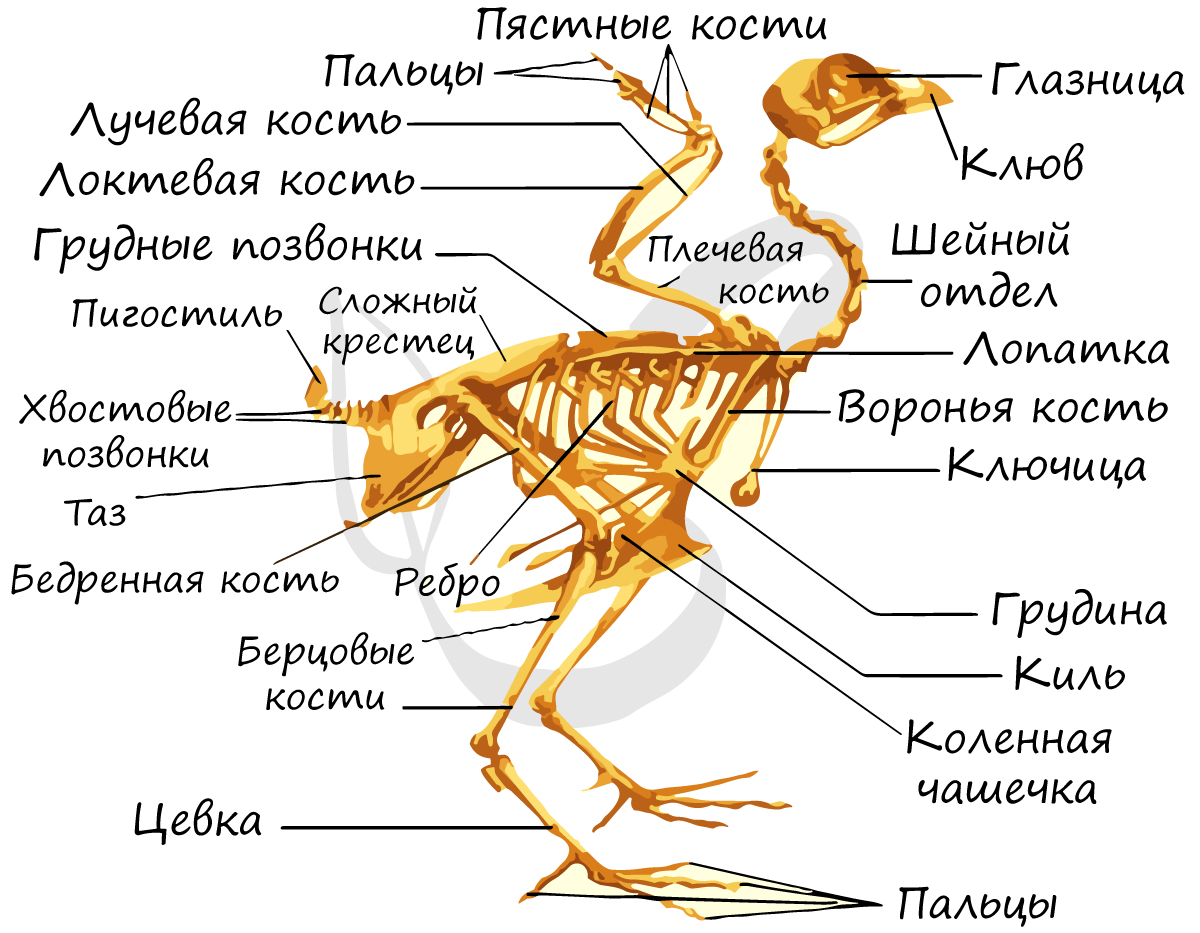
Скелет птиц представлен позвоночным столбом, в состав которого (как и у рептилий) входит 5 отделов: шейный, грудной,
поясничный, крестцовый и хвостовой. Исключительной подвижностью отличается шейный отдел: например, совы могут поворачивать
голову до 270°, что отлично позволяет им выслеживать жертву.

Особенностями скелета является наличие киля — костного выроста грудины, к которому прикрепляются хорошо развитые грудные мышцы,
участвующие в полете. Многие части скелета у птиц срастаются (уменьшается их количество), за счет чего общая масса птицы уменьшается: грудные позвонки сливаются в спинную кость, а хвостовые — в сложный крестец. Срастаются также парные кости тазового пояса.
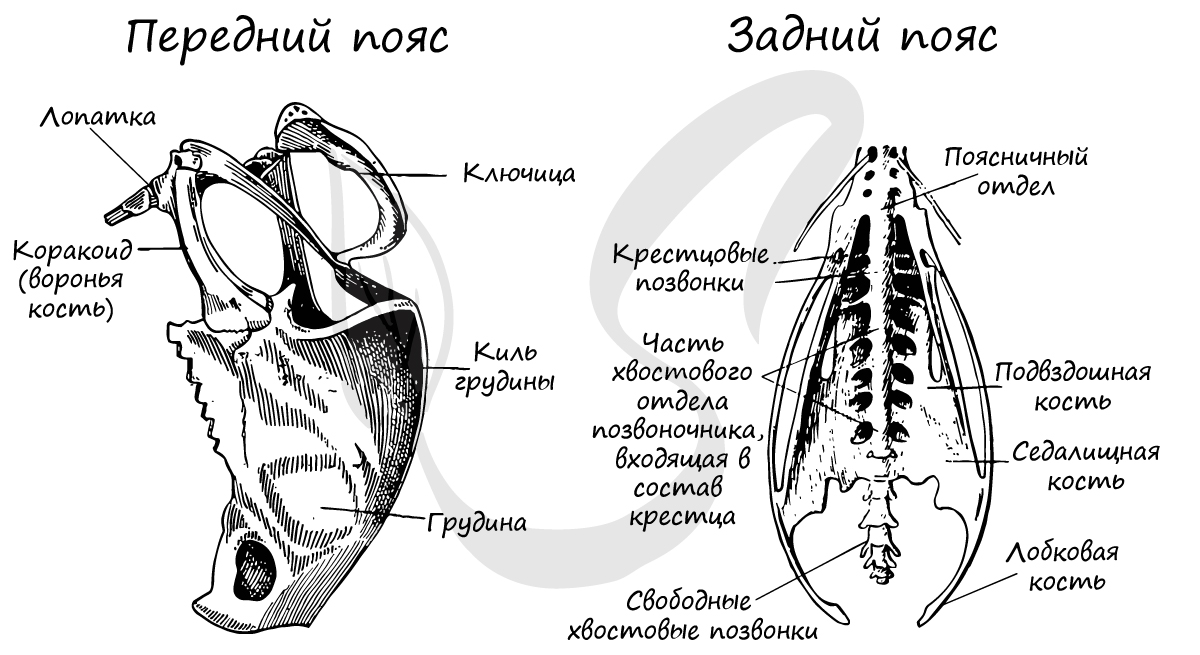
Пояс передних конечностей (плечевой) образован тремя парными костями: вороньими (коракоидами), ключицами и лопатками. Сросшиеся концы ключиц образуют вилочку, которая амортизирует толчки при взмахах крыла.
Тазовый (задний) пояс птиц образован сросшимися друг с другом лобковыми, седалищными, подвздошными костями с каждой стороны. Важно заметить:
лобковые кости не срастаются между собой, что делает таз птиц «открытым», позволяет откладывать крупные яйца.
В ходе приспособления к полету у птиц многие кости становятся полыми внутри. Их можно представить в виде полого цилиндра,
заполненного воздухом, что уменьшает общий вес птицы.
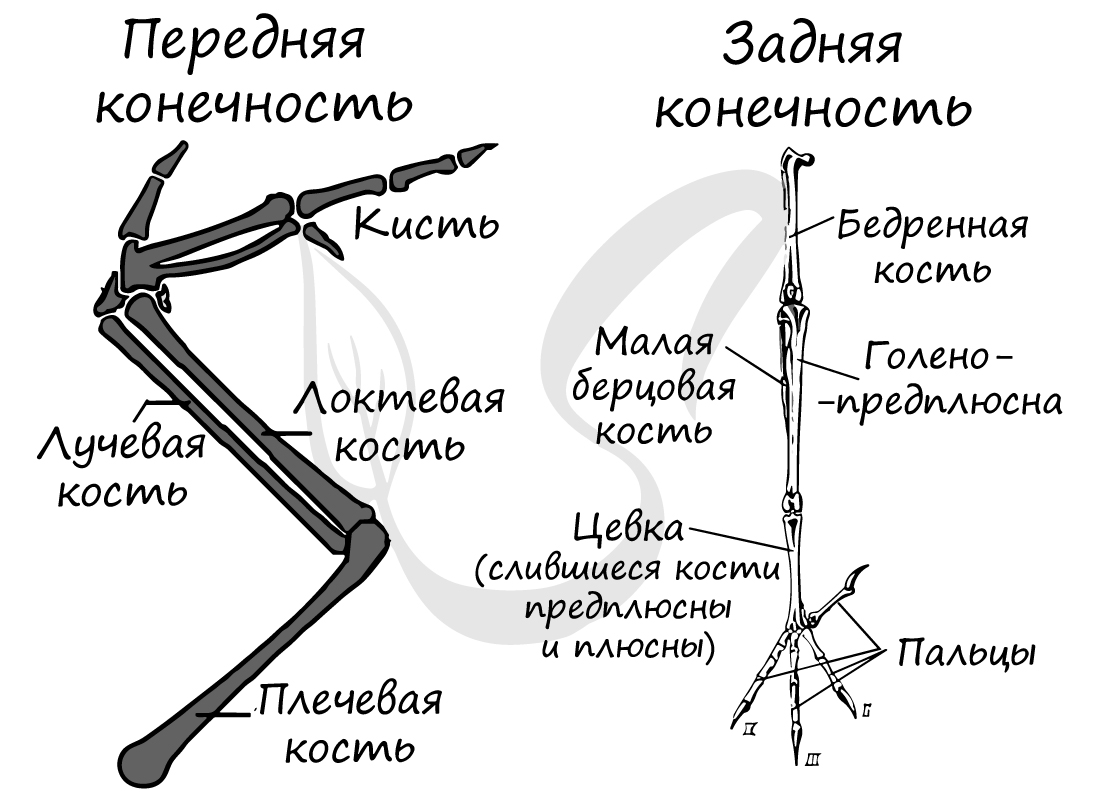
Скелет свободной передней конечности (крыла) состоит из плечевой кости (плечо), лучевой и локтевой кости (вместе формируют предплечье) и кисти (запястье,
пястье, состоящее из одной кости — пряжки, фаланги пальцев). Задняя конечность образована бедренной костью (бедро), малой и большой
берцовыми костями (срастаются друг с другом, образуют голень) и стопы (плюсна, фаланги пальцев).
Особо отметим, что стопа состоит из двух отделов — предплюсна отсутствует, так как срастается с голенью и плюсной. В результате такого
сращение образуется длинная кость — цевка, которая служит амортизатором при приземлении птицы, а также помогает отталкиваться от
земли при взлете, удерживаться на ветке.
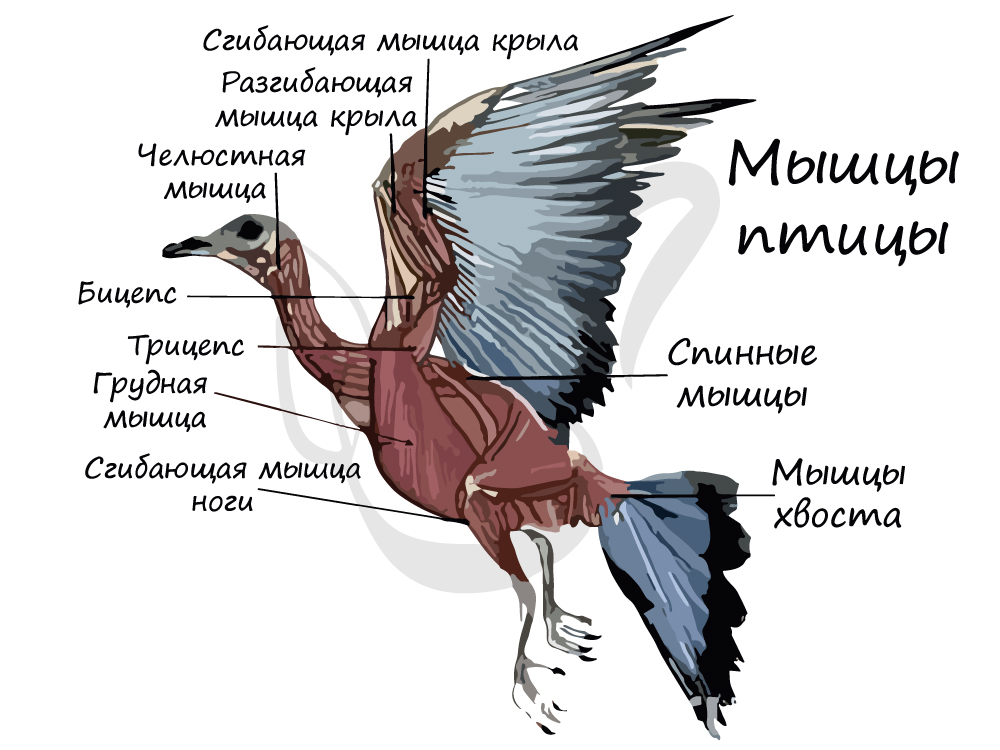
Наиболее развиты у птиц грудные мышцы — опускающие крылья, которые преодолевают большое сопротивление, помогая удерживать птицу в воздухе.
Подключичные мышцы, участвующие в поднимании крыльев также хорошо развиты. В целом мышечная система птиц более сложная и дифференцированная
по сравнению с мышечной системой их предков.
Для добывания пищи и ее захвата птице служит клюв — орган, образованный удлиненными беззубыми челюстями, одетыми роговым
чехлом с режущим краем. Зубы у птиц отсутствуют, однако встречаются самые разные формы клювов, отражающие пищевую специализацию птиц.
Вы уже знаете, что загнутый на конце клюва крючок характерен для хищных птиц: он помогает удерживать жертву, разрывать добычу
на части. У птиц питающихся семенами (воробьиные) небольшой крепкий клюв, снабжен режущими краями, которые позволяют разгрызать оболочки семян.
Удлиненные формы клювов характерны для птиц, охотящихся в воде — цапли, питающихся нектаром — колибри.
Для гусеобразных водоплавающих птиц (гуси, лебеди, утки) характерно наличие плоского клюва, по краю которого расположены роговые пластинки — цедильный аппарат.

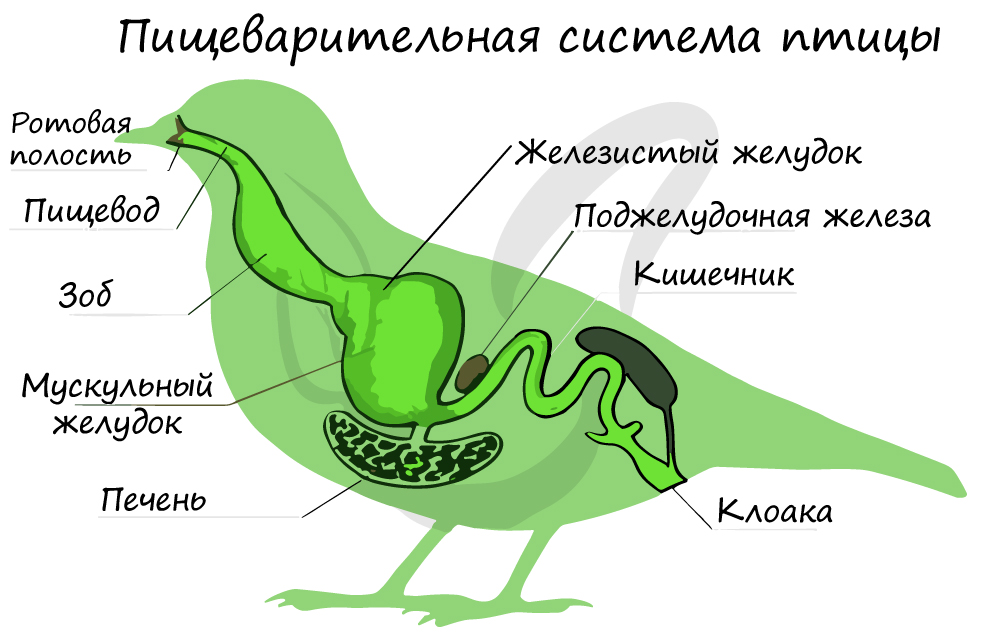
Пищеварительный тракт начинается ротовой полостью, в которой расположен язык. Сюда же открываются протоки слюнных желез.
Ротовая полость переходит в глотку, которая продолжается в пищевод, по ходу которого имеется расширение — зоб. Зоб
предназначен для накапливания и размягчения пищи.
Далее располагается желудок, состоящий из двух отделов: железистого и мускульного. В железистом отделе происходит ферментативная
(химическая) обработка пищи. Мускульный отдел изнутри выстлан плотным роговым покровом, который осуществляет механическую обработку
пищи, измельчая ее.
Желудок продолжается в тонкую кишку, которая сразу открывается в клоаку. Таким образом «полет» оставил след и в этой системе:
пищеварительная трубка птиц укорочена для того, чтобы быстрее и эффективнее расщеплять пищу. Надо отметить, что полет
требует затраты большого количества энергии.
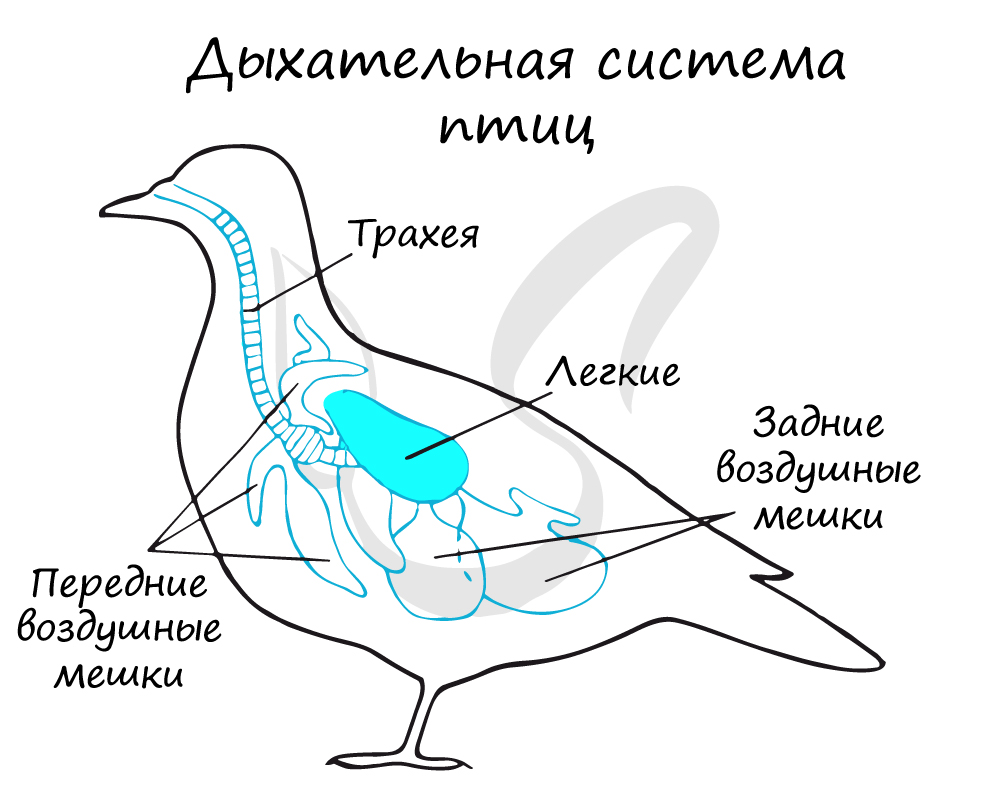
Состоит из ноздрей, ведущих в носовую полость, которая переходит в ротовую, а затем — в трахею. Трахея делится на
два бронха, каждый из которых входит в соответствующее легкое и распадается на бронхи более мелкого калибра.
Легкие птиц отличаются от легких земноводных и рептилий — они имеют губчатое строение, их дыхательная поверхность гораздо
больше. Кроме того, тип дыхания птиц отличается от других наземных животных — им свойственно двойное дыхание, которое
становится возможным благодаря воздушным мешкам.
Воздушные мешки — выросты стенок бронхов, представляющие собой тонкостенные, заполняющиеся воздухом полости. Смысл двойного дыхания
птиц заключается в том, что газообмен в легких осуществляется и на вдохе, и на выдохе.
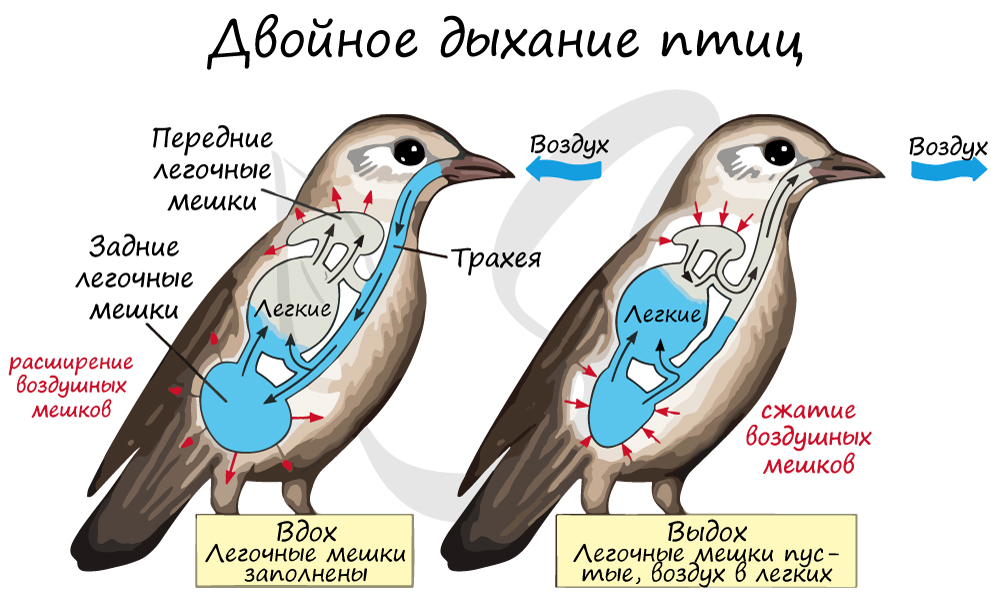
Как возможно, чтобы газообмен шел на выдохе? — спросите вы. На самом деле все просто: во время выдоха воздух выходит из задних
воздушных мешков, и, попадая в легкие, отдает крови кислород, а сам насыщается углекислым газом. Все это ради полета — крайне
сложного двигательного акта, в ходе которого тканям и органам требуется много кислорода.
Кровеносная система птиц замкнутого типа. Сердце с полной перегородкой в желудочке, состоит из 4
камер: двух желудочков и двух предсердий. В левой части сердца кровь артериальная, в правой — венозная. Два круга кровообращения
(большой и малый) оказываются полностью отделены друг от друга.
Обособленность двух кругов кровообращения выводит уровень обмена веществ на самую высокую ступень: птицы становятся теплокровными (в отличие от своих
предков — холоднокровных рептилий). Температура тела птиц более не зависит от окружающей среды, что дает им большие преимущества и
позволяет заселить зоны холодного климата.
Полет крайне энергозатратное удовольствие для кровеносной системы: во время полета сердце птиц развивает
частоту ударов до 1000 в минуту и более, чтобы обеспечить циркуляцию крови адекватную потребностям организма.
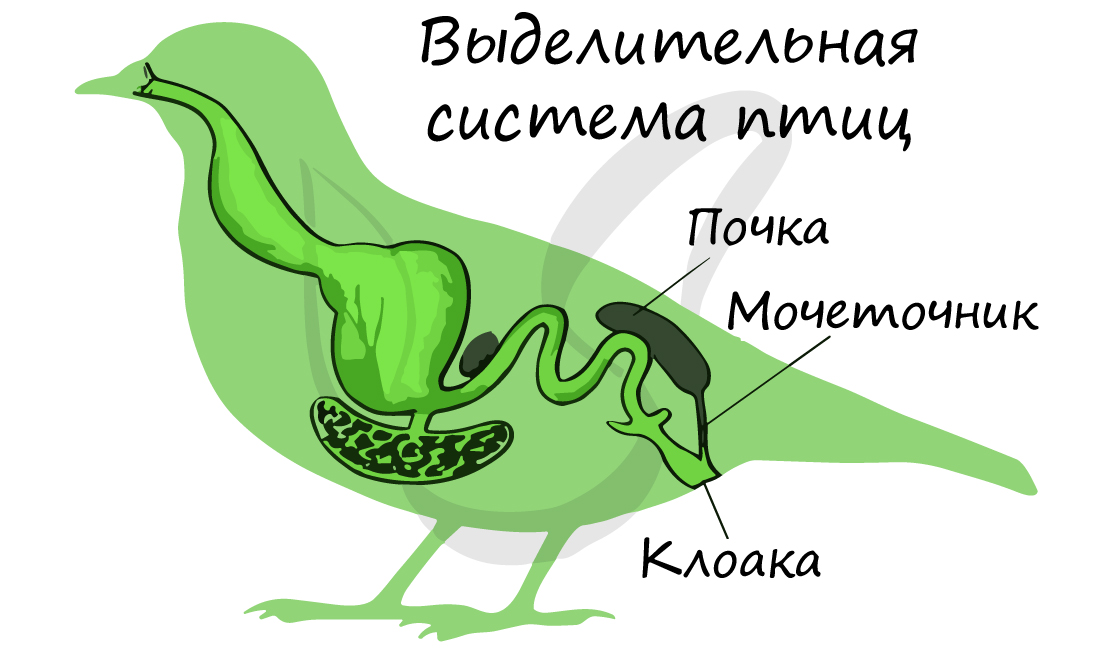
Органы выделения птиц — парные тазовые почки (метанефрос, вторичная почка), от которых начинаются мочеточники, открывающиеся в клоаку. Мочевой пузырь
отсутствует: моча не задерживается в организме, что снижает общий вес птицы. Вследствие отсутствия мочевого пузыря для птиц
характерно частое опорожнение клоаки.
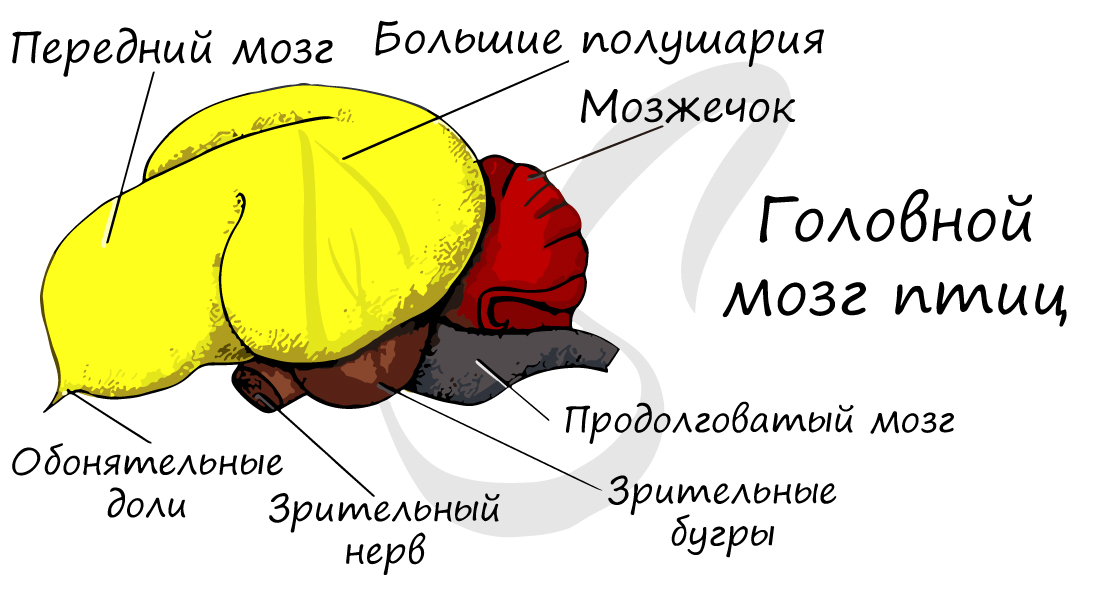
Как и у всех хордовых, нервная система птиц — трубчатого типа. Сравнивая головной мозг птицы и рептилии можно отметить,
что у птиц относительные размеры головного мозга увеличиваются: масса возрастает до 5-8% от массы тела.
Это увеличение происходит за счет развития больших полушарий переднего мозга, которые отвечают за поведенческие реакции. Благодаря этому у птиц наблюдается сложное поведение, выражена забота о потомстве.
У птиц хорошо развит мозжечок, отвечающий за координацию движений, что очень важно для полета. Мозжечок птиц огромный,
складчатый.
Обонятельные доли слабо развиты: бо́льшую роль у птиц играют не органы обоняния, а зрения. Зрительные доли — характерные
образования среднего мозга, представляющие собой вздутия крыши данного отдела. Эти доли хорошо развиты, птицы обладают
исключительным зрением: так, к примеру, орел может заметить мышь в поле на расстоянии 1-2 км.
Птицы практически лишены обоняния, поэтому орган зрения берет на себя основную функцию. Глаз птицы видит острее глаза
человека, к тому же глаза анатомически расположены по бокам головы: такая локализация обеспечивает больший угол обзора.
Строение глаза птиц поражает! Его особенностью является чрезвычайно подвижный хрусталик. Двойной аккомодацией у птиц называется способность хрусталика не только перемещаться вперед-назад, но и менять свою кривизну, что обеспечивают отличную настройку
на наилучшее видение. Даже роговица глаза птицы способна в некоторой степени менять свою кривизну.
Орган слуха развит хорошо, включает внутреннее и среднее ухо. Развитый орган слуха дополняет способность птиц издавать
звуки: так они обмениваются важной информацией друг с другом. Голосовые связки, орган голосообразования, находятся в
нижней гортани (певчей).

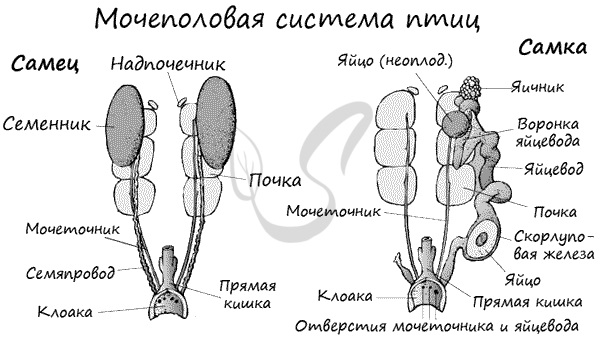
Птицы раздельнополые животные. Мужские половые железы представлены парными семенниками, от которых начинаются семяпроводы.
У голубей перед впадением в клоаку семяпроводы образуют расширение — семенные пузырьки. Специальные копулятивные органы
отсутствуют.
Женская половая система представлена единственным яичником (второй редуцирован для облегчения веса), отходящими от них
яйцеводам, которые открываются в клоаку.
Оплодотворение внутреннее — происходит внутри материнского организма, в яйцеводе.

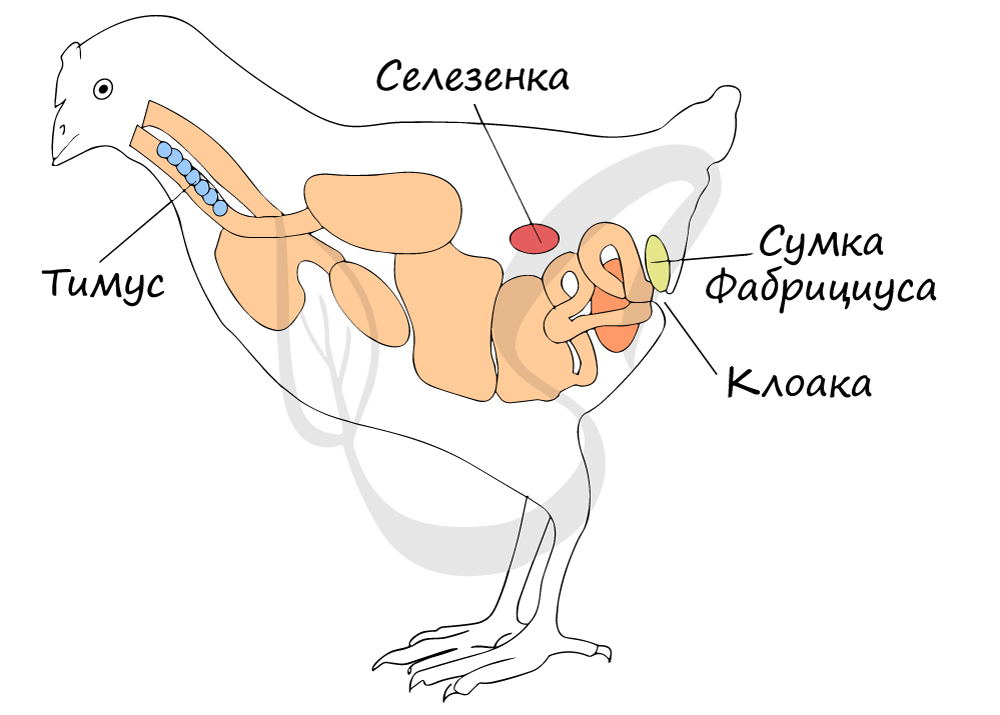
Для всех птиц характерен особый орган — сумка Фабрициуса, открывающаяся протоком в нижнюю
часть клоаки со стороны спины. Фабрициева сумка, как и тимус, — орган иммунной защиты птиц.
Бескилевые птицы
Представители этого отряда характеризуются слабо развитыми крыльями, вследствие чего они неспособны к полету. Этот факт привел к развитию у них выраженной способности к бегу. Например, страусы могут развивать скорость до 70 км/час.
В самом названии отряда заложена главная подсказка: у этих птиц отсутствует киль, костный вырост грудины. В связи с
отсутствием способности к полету их кости не имеют воздухоносных полостей. К данному отряду относятся страусы и
новозеландский киви.

Пингвины
Пингвины — немногочисленная группа птиц, преимущественно обитающих на побережье Антарктиды в Южном полушарии. Способность
к полету у них отсутствует, однако они отлично плавают: их передние конечности превратились в ласты. На грудине имеется киль.
Передвигаются пингвины, опираясь на хвост, не строят гнезд. Крупные виды пингвинов держат яйца на перепонках лап,
пряча под большую складку кожи на животе, насиживают стоя.

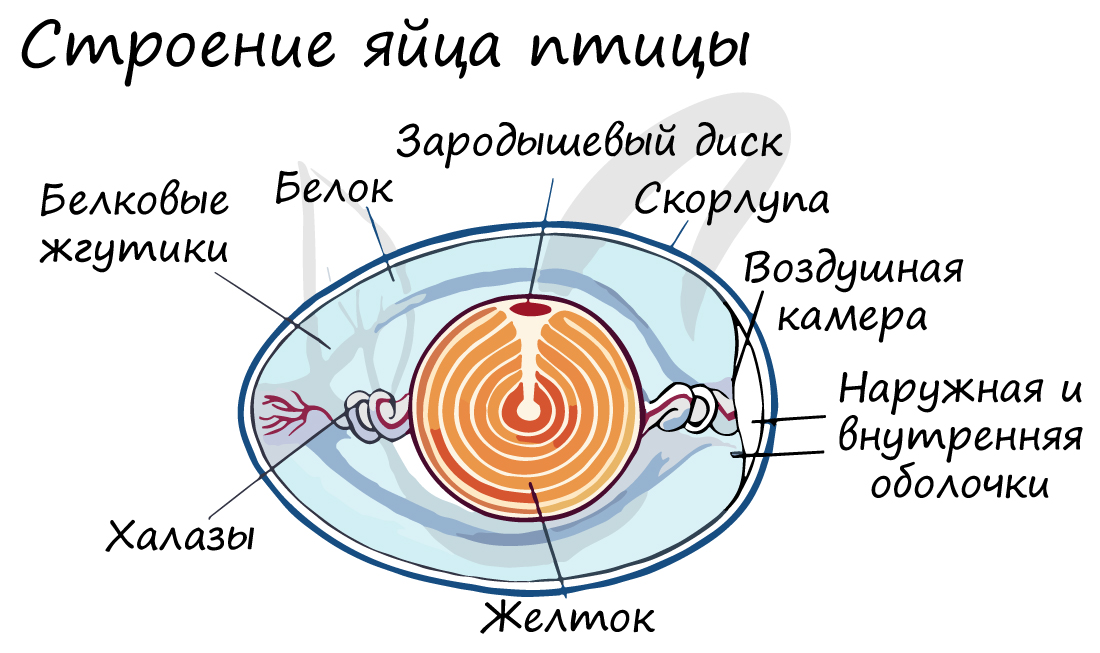
Строение яйца птицы
По мере прохождения оплодотворенного яйца по яйцеводу оно покрывается белковой, волокнистой и скорлуповыми оболочками, которые
надежно его защищают. Собственно яйцом является желток — запас питательных веществ для зародыша, представленного
зародышевым диском.
Желток фиксируется в яйце белковыми жгутиками (нитями), которые, скручиваясь, образуют халазы — канатики из белка. Халазы
подвешивают желток в яйце, предохраняют его от механических повреждений и обеспечивают нужное положение, при котором
зародышевый диск всегда находится сверху.
Белок в яйце выполняет защитную и запасающую функции, обеспечивает зародыш водой. В яйце имеется воздушная (дыхательная)
камера, в которой происходит газообмен с окружающей средой — без газообмена дыхание зародыша остановится, он погибнет.
Птицы откладывают яйца с плотными защитными оболочками, в которых развивается зародыш. Чтобы птенец успешно вылупился,
нужна повышенная температура (около 37-38°C) — самка принимается насиживать яйца. Высиживание занимает около 6 недель.

Выводковые и гнездовые птицы
Настает счастливый момент начала новой жизни — рождение птенца. Однако при множественных наблюдениях легко заметить, что у
одних птиц рождаются птенцы, которые уже через несколько часов самостоятельно могут ходить, летать — такие птицы называются
выводковые.
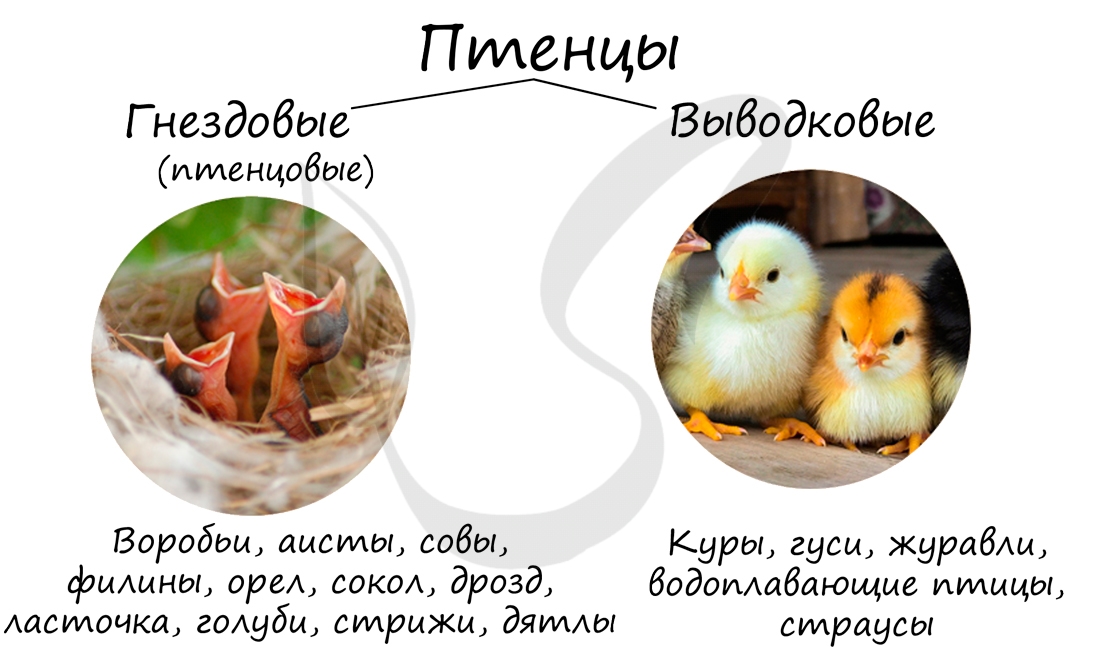
Птенцы выводковых птиц после вылупления покрыты пухом, хорошо слышат. К выводковым птицам относятся гусеобразные, курообразные,
дрофы, журавли, водоплавающие птицы — утки, пингвины.

У других птиц птенцы слепые, глухие, не умеющие ходить: они нуждаются в тщательной защите и выкармливании. Такие птицы называются
гнездовые. Их птенцы голые или слабоопушенные. К ним относятся воробьи, дятлы, голуби, попугаи, кукушки, стрижи и колибри.

Птицы и их образ жизни
По образу жизни птицы подразделяются на оседлых, кочующих и перелетных.
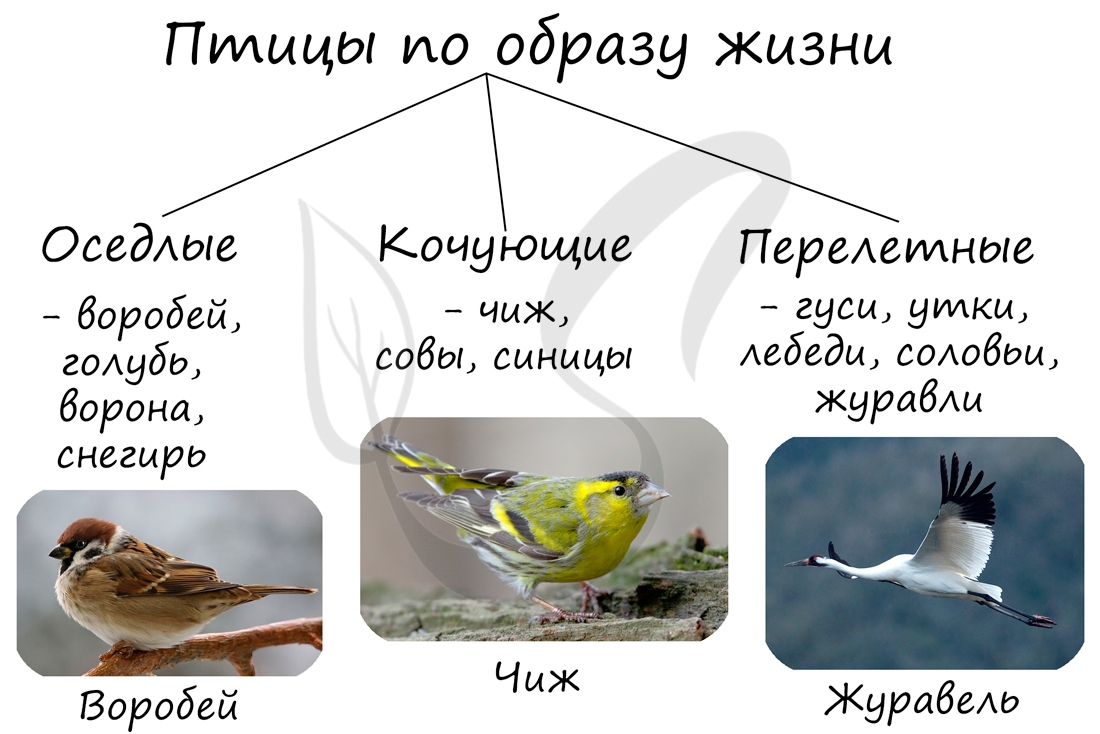
Оседлые птицы живут в пределах небольшой территории и редко покидают ее пределы. Такой образ жизни птиц характерен в местах обитания,
где сезонные изменения не затрагивают доступность корма: в тропических и субтропических зонах.
Помните, что для птиц не страшна низкая температура зимой — они теплокровные! Гораздо важнее доступность корма.

Кочующими птицами называют тех, которые постоянно перемещаются с места на место в поисках пищи вне сезона размножения.
Они могут совершать перелеты на сотни километров, но обычно не покидают той природной зоны, в которой гнездятся.

Перелетные птицы совершают регулярные сезонные миграции между местами гнездовий и зимовок. Они преодолевают тысячи
километров.

Значение птиц
Птицы, как и все живые существа, являются звеном в цепи питания (консументы). Питаются насекомыми, регулируя их численность.
Хищные птицы — санитары леса, питаются ослабленными и больными животными. Имеют важное эстетическое значение: их внешний
вид впечатляет, а раннее утреннее пение приносит нам порой истинное удовольствие
С древних времен люди занимаются птицеводством, дающим множество продуктов, без которых трудно представить наше существование. Мясо домашних птиц богато полезными веществами, питательно, как и куриные яйца. В настоящее время процессы
разведения птиц механизированы: их выращивают в специальных инкубаторах.

© Беллевич Юрий Сергеевич 2018-2023
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
Карташев Н.Н., Соколов В.Е., Шилов И.А.
Практикум по зоологии позвоночных
М., Высшая школа. 1981 г.
ТЕМА 17. СКЕЛЕТ ПТИЦЫ
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ ОБЪЕКТА
Подтип Позвоночные, Vertebrata
Класс Птицы, Aves
Отряд Воробьинообразные, Passeriformes
Представитель — Серая ворона, Corvus corone L.
МАТЕРИАЛ И ОБОРУДОВАНИЕ
Для занятий важно иметь хотя бы один смонтированный скелет птицы. На одного-двух студентов необходимы:
1. Разборный скелет вороны — 1 комплект.
2. Препарировальные иглы — 2.
ЗАДАНИЕ
Уяснить особенности строения скелета птиц. Сделать следующие рисунки:
1. Череп вороны сбоку.
2. Череп вороны снизу.
3. Скелет туловища сбоку (включая позвоночный столб, пояс передних конечностей, грудину, ребра, тазовый пояс).
4. Тазовый пояс и сложный крестец снизу.
5. Скелет крыла.
6. Скелет задней конечности.
7. Первый, второй и один из задних шейных позвонков (вид сбоку).
ОПИСАНИЕ СКЕЛЕТА
В скелете птиц отчетливо выражены черты, связанные с приспособлением к полету. Плоские кости обычно очень тонкие, губчатые. В больших трубчатых костях хорошо развиты полости, заполненные или костным мозгом, или воздухом. Все это обеспечивает повышенную прочность скелета и заметно облегчает его. Нужно, однако, иметь в виду, что общая масса скелета птиц составляет 8—18% от массы тела, т. е. примерно столько же, сколько и у млекопитающих (6—14% веса тела), хотя у последних кости толще, а воздушные полости в них отсутствуют. Это объясняется тем, что у птиц значительно больше относительная длина скелетных элементов конечностей и некоторых других костей.
Скелет птиц подразделяется на осевой скелет, грудную клетку (грудина и ребра), череп, скелет конечностей и их поясов.
Осевой скелет. Осевой скелет, или позвоночный столб (column. vertebralis), как и у пресмыкающихся, разделяется на пять отделов шейный, грудной, поясничный, крестцовый и хвостовой.
Шейный отдел очень подвижен (птицы легко поворачивают голов на 180°, некоторые даже на 270°). Число шейных позвонков варьирует в широких пределах: от 11 (попугаи) до 23—25 (лебеди, утки); у вороны, голубя 14 шейных позвонков.

Рис. 105. Шейные позвонки вороны
А — атлант; Б — эпистрофей; В — средний шейный позвонок:
1 — сочленовная ямка для затылочного мыщелка, 2 —канал для спинного мозга, 3 — тело позвонка, 4 — зубовидный отросток, 5 — верхняя дуга, 6 — остистый отросток, 7 — передние сочленовные отростки, 8 — задние сочленовные отростки, 9 — поперечный отросток, 10 — шейное ребро, 11 — позвоночное отверстие
Первый шейный позвонок — атлас, или атлант (atlas; рис. 105, А) имеет форму кольца. На нижней части его передней поверхности есть глубокая сочленовная ямка, в которую входит затылочный мыщелок черепа; на нижней задней поверхности — сочленовная плоскость для соединения с телом второго шейного позвонка. Просвет полости кольца поперечной сухожильной, обычно окостеневающей связкой разделен на две части: через верхнюю проходит спинной мозг, а в нижней помещается зубовидный отросток второго позвонка.
Второй шейный позвонок — эпистрофей (epistrophaeus; рис. 105, Б) имеет хорошо развитое тело с направленным вперед зубовидным отростком (рис. 105, 4). По происхождению зубовидный отросток, видимо, — тело первого позвонка, обособившееся от него и сросшееся с телом второго позвонка. Сбоку расположены поперечные отростки; к ним крепятся шейные мышцы, соединяющие второй позвонок с затылочным отделом черепа и с другими шейными позвонками. Над телом позвонка расположена хорошо развитая верхняя дуга (рис. 105, 5), через просвет которой проходит спинной мозг.
Остальные шейные позвонки имеют гетероцельное строение: довольно длинное тело каждого позвонка (corpus vertebrae; рис. 105, 3) на передней и задней стороне имеет седловидные сочленовные поверхности. Такой характер сочленения обеспечивает значительную подвижность позвонков друг относительно друга, как в вертикальной, Так и в горизонтальной плоскостях. Это обеспечивает наряду со сложной системой дифференцированных шейных мышц большую подвижность шеи, что очень важно для птиц (компенсация неподвижного скелета туловища, возможность менять положение центра тяжести в полете, сгибая и выпрямляя шею, возможность сложных движений головы при чистке оперения, постройке гнезда и т. п.).
Над телом позвонка расположена верхняя дуга (arcus neuralis; рис. 105, 5), заканчивающаяся остистым отростком (processus spinosus; рис. 105, 6); в канале дуги проходит спинной мозг. От передней и задней поверхности дуги отходят парные сочленовные отростки (рис. 105, 7, 8), соединяющиеся с такими же отростками соседних позвонков. Такой тип сочленения обеспечивает прочность соединения позвонков друг с другом при сохранении значительной подвижности. От боковой поверхности тел позвонков отходят короткие поперечные отростки (processus transversus; рис. 105, 5). Птицы, как и пресмыкающиеся, имеют шейные ребра (рис. 105, 10); они рудиментарны и только последние одно-два шейных ребра достаточно длинны и подвижны (рис. 106, 5), но не доходят до грудины. Шейное ребро прирастает своей головкой к телу позвонка, а бугорком — к поперечному отростку. Таким путем образуются позвоночные отверстия (foramen transversarium; рис. 105, 1), создающие с обеих сторон шеи канал, по которому проходят позвоночная артерия и шейный симпатический нерв. Эта костная защита артерии и нерва особенно важна при длинной, очень подвижной шее.

Рис. 106. Скелет туловища вороны сбоку:
1 — последние шейные позвонки, 2 — сросшиеся грудные позвонки (спинная кость), 3 — шейные ребра, 4 — грудные ребра, 5 — крючковидный отросток, 6 — тело грудины, 7 — киль, грудины, 8 — сложный крестец, 9 — подвижные хвостовые позвонки, 10 — пигостиль, 11 — коракоид, 12 — вилочка, 13 — лопатка, 14 — суставная впадина для головки плеча, 15 — подвздошная кость,
16 — седалищная кость, 17 — лобковая кость, 18 — вертлужная впадина для сочленения с головкой бедра
Грудной отдел у вороны представлен шестью позвонками (у других видов птиц их число варьирует от 3 до 10), которые, как и у большинства птиц, срастаются друг с другом, образуя спинную кость (dorsale; рис. 106, 2).
К грудным позвонкам подвижно причленяются грудные ребра (costa; рис. 106, 4); число их пар равно числу грудных позвонков. Это плоские, слегка изогнутые костные пластинки, состоящие из двух подвижно соединенных друг с другом отделов — спинного и брюшного. Верхний конец спинного отдела ребра имеет два сочленовных отростка: бугорок, причленяющийся к поперечному отростку, и головку, сочленяющуюся с телом позвонка. Нижний конец брюшного отдела ребра подвижно сочленяется с телом грудины (sternum рис. 106, 6). Подвижное соединение ребер с грудиной и позвоночным столбом вместе с подвижным сочленением спинного и брюшного отделов ребра обеспечивает возможность значительных изменений объема полости тела путем опускания грудины вниз и поднимания ее кверху. Эта подвижность возможна благодаря сильно развитой реберной мускулатуре. Жесткость грудной клетки усиливается костными образованиями — крючковидными отростками (processus uncinatus; рис. 106, 5) ребер, укрепляющимися на спинном отделе и налегающими на последующее ребро.
Грудина, или грудная кость (sternum; рис. 106, 6) — широкая и длинная, вогнутая изнутри костная пластинка, несущая по средней линии высокий костный гребень — киль грудины (crista sterni; рис. 106, 7). Резкое увеличение поверхности грудины, обусловленное как ее разрастанием, так и образованием киля, — прямое приспособление к полету; оно обеспечивает возможность прикрепления мощных мышц, приводящих в движение крыло. На переднем конце грудины по бокам киля расположены большие сочленовные площадки, обеспечивающие прочное прикрепление к телу грудины коракоидов (рис. 106, 11). На боковых краях грудины размещаются мелкие углубления, которые служат для сочленения с нижними концами брюшных отделов ребер.

Рис. 107. Сложный крестец вороны (вид снизу):
1 — поясничные позвонки, 2 — крестцовые позвонки, 3 — хвостовые позвонки,
4 — подвздошная кость, 5 — седалищная кость, 6 — лобковая кость
За грудным отделом позвоночного столба расположен сложный крестец (synsacrum; рис. 106, 8; рис. 107), представляющий собой слившиеся в общую монолитную кость все поясничные (у ворон и голубей 6), все крестцовые (почти у всех видов птиц 2) и часть хвостовых (3—8) позвонков. Сложный крестец неподвижно срастается с последним грудным позвонком. Границы между отделами в сложном крестце провести трудно; крестцовые позвонки (рис. 107, 2) бывают заметны лишь по отверстиям между их поперечными отростками и подвздошными костями. Кости тазового пояса также неподвижно срастаются с позвонками сложного крестца.
Благодаря срастаниям позвонков туловищный участок осевого скелета птиц представляет собой прочное, монолитное образование, обеспечивающее неподвижность туловища, что крайне важно при полете. Образование сложного крестца, слившегося с костями тазового пояса, создает прочную опору для задних конечностей, на которые давит вся тяжесть тела при движении птицы по земле (рис. 107).
Число свободных, подвижно соединенных друг с другом хвостовых позвонков (рис. 106, 9) у птиц невелико (от 5 до 9; у вороны и голубя — обычно 7). Последние хвостовые позвонки (4—8) сливаются в вертикально уплощенную пластинку — копчиковую кость (pygostyle; рис. 106, 10), к боковым поверхностям которой веером прикрепляются основания рулевых перьев. Укорочение хвостового скелета увеличивает общую компактность тела (важно аэродинамически). Характер прикрепления рулевых перьев позволяет использовать хвост в полете (добавочная несущая плоскость) и при посадке (торможение).
Череп. Череп птиц тропибазального типа (мозг не лежит между глазницами). По сравнению с пресмыкающимися резко бросается в глаза значительное увеличение объема мозговой полости, большие глазницы, появление клюва, полное исчезновение зубов, у большинства птиц — заметное смещение вниз большого затылочного отверстия. Кости черепа настолько тонки, что соединение их друг с другом при помощи швов становится невозможным. Поэтому у птиц большинство костей черепа сливается друг с другом; границы между отдельными костями хорошо видны только на черепе птенцов. Тонкие кости и редукция зубов сильно облегчают скелет головы. В то же время благодаря сложной губчатой внутренней структуре кости черепа сохраняют большую прочность.
Череп (cranium) делится на мозговой череп (neurocranium), включающий собственно мозговую (черепную) коробку, носовой отдел и область глазниц, и висцеральный череп (splanchnocranium), к которому относятся челюсти (клюв), кости нёба и подъязычный аппарат.

Рис. 108. Череп вороны
А — сбоку; Б — снизу; В — сверху:
1 — большое затылочное отверстие, 2 — основная затылочная кость, 3 — боковая затылочная кость, 4 — верхняя затылочная кость, 5 — затылочный мыщелок, 6 — теменная кость, 7 —лобная кость, 8 — носовая кость, 9 — чешуйчатая кость, 10 — наружный слуховой проход, 11 — боковая клиновидная кость, 12 — средняя обонятельная кость, 13 — предлобная кость, 14 — предчелюстная кость, 15 — верхнечелюстная кость, 16 — скуловая кость, 17 — квадратно-скуловая кость, 18 — квадратная кость, 19 — основная височная кость, 20 — клювовидный отросток парасфеноида, 21 — сошник, 22 — нёбная кость, 23 — крыловидная кость, 24 — зубная кость, 25 — сочленовная кость, 26 — угловая кость
Начнем рассмотрение черепа с затылочной области. На задне-нижней стенке черепной коробки находится большое затылочное отверстие (foramen occipitale magnum; рис. 108, 1), окруженное четырьмя сросшимися затылочными костями. Вентрально расположена основная затылочная кость (basioccipitale; рис. 108, 2), по бокам — парные боковые затылочные кости (occipitale laterale или exooccipitale; рис. 108, 3). Основная и боковые затылочные кости образуют единственный, как и у пресмыкающихся, затылочный мыщелок (condylus occipitalis; рис. 108,5), сочленяющий череп с первым шейным позвонком. Над затылочным отверстием располагается верхняя затылочная кость (supraoccipitale; рис. 108, 4).
Спереди от верхней и боковых затылочных костей лежат парные теменные кости (parietale; рис. 108, 6). Крышу черепа над глазницами, верхние и задние стенки глазниц и всю переднюю часть мозговой коробки образуют длинные и широкие лобные кости (frontale; рис. 108, 7). Впереди лобных костей у основания клюва лежат парные носовые кости (nasale; рис. 108, с двумя отростками: верхний идет вперед сбоку от гребня клюва и ограничивает носовое отверстие, сверху, челюстной — вперед и вниз к верхнечелюстной кости и ограничивает носовое отверстие сзади. Боковую часть мозговой коробки образуют парные крупные чешуйчатые кости (squamosum; рис. 108, 5), лежащие книзу от лобных и спереди от теменных и боковых затылочных костей. Верхняя височная дуга у птиц отсутствует. Чешуйчатая кость прикрывает ушные кости, которые срастаются вместе и образуют костные стенки среднего и внутреннего уха; снаружи они обычно не видны. При хорошей препарировке черепа в центре ушного прохода (рис. 108, 10) бывает виден конец тонкой столбчатой косточки (или стремечка, stapes), образовавшийся в результате окостенения подвеска (hyomandibulare) — верхнего элемента подъязычной дуги.
Ниже лобных костей переднюю часть мозговой коробки (являющуюся также задней стенкой глазницы) образуют небольшие парные боковые клиновидные кости (laterosphenoidem; рис. 108, 11). Практически почти всю тонкую межглазничную перегородку образует непарная средняя обонятельная кость (mesethmoideum; рис. 108, 12). Передние стенки глазницы образуют парные (по одной с каждой стороны) предлобные кости (praefrontale; рис. 108, 13); у вороны и других воробьиных птиц они очень малы, и основную часть передней стенки глазницы заполняет отросток средней обонятельной кости.
Клюв птицы состоит из двух частей: надклювья, образованного предчелюстными, верхнечелюстными и носовыми костями, и подклювья. Большую часть надклювья составляют слившиеся парные предчелюстные кости (praemaxillare; рис. 108, 14). Спереди они образуют выпуклую сверху л слегка вогнутую снизу вершину клюва, а назад продолжаются в виде трех парных отростков:
1) лобные отростки соединяются с верхними отростками носовых костей и образуют верхний гребень клюва (ограничивают носовое отверстие сверху и спереди);
2) челюстные отростки отграничивают носовое отверстие сбоку;
3) нёбные отростки образуют костное дно надклювья (они входят в состав твердого нёба).
Небольшие верхнечелюстные кости (maxillare; рис. 108, 15) образуют лишь заднюю нижнюю часть надклювья, соединяясь спереди с челюстными отростками предчелюстных костей; снизу к ним примыкают (сливаясь) челюстные отростки нёбных костей.
От заднего края верхнечелюстной кости отходит назад тонкая костная перекладина, состоящая из двух слившихся костей — скуловой (jugale; рис. 108, 16) и квадратно-скуловой (quadrato-jugale; рис. 108, 17). По своему положению это типичная нижняя дуга, поэтому череп птиц относят к диапсидному типу с редуцированной верхней дугой. Квадратно-скуловая кость причленяется к квадратной кости (quadratum; рис. 108, 18). Нижний конец квадратной кости образует суставную поверхность для причленения нижней челюсти, а другой удлиненный ее конец подвижно сочленяется с чешуйчатой и переднеушной костями.
Посмотрим теперь на череп снизу. В основании черепа, впереди основной затылочной кости (рис. 108, 2) лежит небольшая основная клиновидная кость (basisphenoideum). Она полностью прикрыта широкой основной височной костью (basitemporale; рис. 108, 19) — производной парасфеноида. Передняя часть парасфеноида сохраняется в виде направленного вперед узкого клювовидного отростка (rostrum parasphenoidei; рис. 108, 20). У его переднего конца лежит сошник (vomer; рис. 108, 21). По бокам сошника располагаются хоаны — внутренние отверстия ноздрей.
Нёбные отростки предчелюстных и верхнечелюстных костей сливаются с удлиненными парными нёбными костями (palatinum; рис. 108, 22) и образуют костное дно надклювья. Задние, имеющие сложный профиль, концы нёбных костей налегают на клювовидный отросток парасфеноида. В этом месте к нёбным костям причленяются (суставом) парные крыловидные кости (pterygoideum; рис. 108, 23), задние концы которых также суставом соединяются с квадратными костями.

Рис. 109. Схема кинетизма черепа птицы
Сплошной линией и пунктиром показано изменение положения надклювья при движении костного нёба:
1 — квадратная кость, 2 — крыловидная кость, 3 — нёбная кость, 4 — скуловая кость, 5 — квадратно-скуловая кость, 6 — надклювье, 7 — место перегиба костей
Эти элементы твердого нёба имеют очень важное значение для движения клюва (рис. 109). Если сокращаются мышцы, соединяющие направленный вперед длинный глазничный отросток квадратной кости со стенками глазницы, то направленный вниз конец квадратной кости несколько подвигается вперед и толкает вперед как крыловидные и нёбные кости (место сочленения этих костей может скользить по клювовидному отростку парасфеноида, на который они налегают), так и квадратно- скуловую и скуловую. Давление по этим двум костным мостикам передается на основание надклювья, благодаря чему вершина надклювья приподнимается кверху. При этом прогибаются кости у основания вершины надклювья, в области «переносицы». Перегиб облегчает очень сильное истончение костей в этом месте; у ряда видов здесь сохраняется хрящевая перемычка или даже образуется настоящий сустав. При сокращении мышц, соединяющих череп с нижней челюстью, нижний конец квадратной кости сдвигается назад, оттягивая на себя эти костные связи, и вершина клюва сдвигается книзу.
Описанный сложный костный механизм твердого нёба (основа его — подвижно соединенная с черепом сложной формы квадратная кость) дополняется системой дифференцированных жевательных мышц. Все это обеспечивает возможность довольно разнообразных движений клюва, облегчающих захват добычи, чистку оперения, постройку сложного гнезда и т. п. Возможность дифференцированных движений клюва выработалась, видимо, в связи с преобразованием передних конечностей в крылья, выполняющие только функцию полета.
Подклювье — нижняя челюсть — представляет собой ряд слившихся костей, из которых наиболее крупные зубная (dentale; рис. 108, 24), сочленовная (articulare; рис. 108, 25) и угловая (angulare; рис. 108, 26). Челюстной сустав образуют, соединяясь друг с другом, квадратная и сочленовная кости.

Рис. 110. Подъязычный аппарат вороны:
1 — тело, 2 — его передний отросток, служащий основанием языка, 3 — рожки
Остатки гиоида и жаберных дуг превращаются в подъязычный аппарат (рис. 110). У некоторых птиц (например, дятлов) благодаря большой длине рожков и общей подвижности подъязычного аппарата язык может выдвигаться вперед почти на длину клюва, что облегчает захват добычи.
Конечности и их пояса. Пояс передних конечностей (плечевой пояс) птиц образован парными коракоидами, лопатками и ключицами.
Мощные удлиненные коракоиды (coracoideum; см. рис. 106, 11) своими расширенными нижними концами прочно соединяются малоподвижными суставами с передним краем грудины. Между передними концами коракоидов расположена прикрепляющаяся к ним вилочка (furcula, см. рис. 106, 12), возникшая путем слияния обеих ключиц. Длинные и узкие лопатки (scapula; см. рис. 106, 13) лежат над ребрами; их передние концы прочно соединены со свободными концами коракоидов. В месте соединения лопатки и коракоида расположена довольно глубокая суставная впадина (см. рис. 106, 14), в которую входит головка плечевой кости.
Мощность костей плечевого пояса и их прочное соединение с грудиной обеспечивают опору для крыльев в полете. Удлинение коракоидов увеличивает площадь прикрепления мышц крыла, а также позволяет вынести вперед плечевой сустав, что обеспечивает и удобное складывание крыла в покое, и аэродинамически выгодное его положение в полете, когда центры площадей крыльев лежат на линии, проходящей через центр тяжести птицы. Расположенная между свободными концами коракоидов вилочка обычно играет роль пружины, амортизирующей резкие толчки при взмахах крыла в полете.

Рис. 111. Скелет крыла вороны:
1 — плечевая кость, 2 — лучевая кость, 3 — локтевая кость, 4 —самостоятельные косточки запястья, 5 — пряжка (слившиеся косточки запястья и пясти), 6 — фаланги II пальца, 7 — единственная фаланга I пальца, 8 — единственная фаланга III пальца
Скелет передней конечности, превратившейся в крыло, подвергся значительным изменениям. Плечо (humerus; рис. 111, 1) — мощная трубчатая кость, проксимальный конец которой образует уплощенную головку плеча, входящую в суставную впадину плечевого сустава. Характер суставных поверхностей заметно ограничивает возможность вращательных движений в плечевом суставе, что обеспечивает устойчивость крыла в полете. Мощные эпифизы дистального конца плеча образуют суставные поверхности для сочленения с двумя костями предплечья: более прямой и тонкой лучевой (radius; рис. 111, 2) и более мощной, слегка изогнутой локтевой костью (ulna; рис. 111, 3). На поверхности локтевой кости видны бугорки — места прикрепления очинов второстепенных маховых перьев. Характер суставных поверхностей локтевого сустава обеспечивает прочное соединение костей плеча и предплечья и ограничивает вращательные движения и в этом суставе. При этом сохраняется большая подвижность в одной плоскости — плоскости крыла, что дает возможность птице складывать крыло в покое и изменять его площадь при изменении режима полета.
Особенно резкие изменения в связи с приспособлением к полету произошли в строении кисти. В проксимальном отделе запястья (carpus) сохраняются только две самостоятельные косточки (рис. 111, 4). Они связками почти неподвижно соединяются с костями предплечья. Подвижный сустав расположен между ними и остальными костями запястья, поэтому его называют интеркарпальным. Остальные кости запястья и все кости пясти (metacarpus) сливаются в единое образование— пряжку (carpometacarpus; рис. 111, 5). Резко редуцируется скелет пальцев. Сохраняются две фаланги II пальца (рис. 111, 6), продолжающие ось пряжки. От I пальца сохраняется одна фаланга (рис. 111, 7), к ней прикрепляется пучок перьев, образующий крылышко (alula; см. рис. 98, 4). III палец представлен также одной фалангой (рис. 111, 8), прикрепленной к основанию первой фаланги II пальца.
Эти преобразования в скелете кисти (образование carpometacarpus, редукция пальцев) обеспечивают прочную опору для первостепенных маховых перьев — части крыла, несущей в полете наибольшую нагрузку.
Тазовый пояс состоит из трех пар срастающихся вместе костей: Широкая и длинная подвздошная кость (ilium; см. рис. 106, 15; рис. 107, 4) на большом протяжении срастается со ложным крестцом. К ее наружному краю прирастает седалищная кость (ischium; см. рис. 106, 16; рис. 107, 5), а ниже расположена направленная назад палочкообразная лобковая кость (pubis; см. рис. 106, 17; рис. 107, 6). Все три кости таза образуют вертлужную впадину (acetabulum; см. рис. 106, 18), в которую входит головка бедра.
Большая поверхность тазовых костей и их прочное (неподвижное) соединение с осевым скелетом обеспечивают прочную опору для прикрепления мощных мышц. Эти особенности тазового пояса определяются тем, что передние конечности выполняют только функцию полета, а при движении по земле вся тяжесть тела поддерживается лишь задними конечностями.
Лобковые и седалищные кости правой и левой сторон не соединяются друг с другом на брюшной поверхности (см. рис. 107). Поэтому таз птиц называют открытым; он дает возможность откладывать крупные яйца. Можно предполагать, что открытый таз, увеличивающий подвижность брюшной стенки в тазовой области, способствует интенсификации дыхания в полете.

Рис. 112. Скелет задней конечности вороны:
1 — бедренная кость, 2 — коленная чашечка, 3 — голено-предплюсна (tibio-tarsus), 4 — малая берцовая кость, 5 — цевка (слившиеся кости предплюсны и плюсны), 6 – интертарзальный сустав, 7 — фаланги пальцев; I — IV — пальцы
Скелет задней конечности представлен мощными трубчатыми костями. Бедро (femur; рис. 112, 1) на проксимальном конце имеет хорошо развитую округлую головку для сочленения с тазовым поясом. На дистальном конце образуются рельефные суставные поверхности для сочленения с костями голени. В области коленного сустава в мышечном сухожилии лежит коленная чашечка (patella; рис. 112, 2).
Основной элемент голени — массивная большая берцовая кость, с нижним дистальным концом которой сливаются две проксимальные кости предплюсны; образуется костный комплекс, представляющий собой голено-предплюсну, или тибио-тарзус (tibio-tarsus; рис. 112, 3). Малая берцовая кость (fibula; рис. 112, 4) сильно редуцирована и в виде маленькой тоненькой косточки прирастает к верхней части наружной поверхности большой берцовой кости.
Дистальные элементы предплюсны (tarsus) и все элементы плюсны (metatarsus) сливаются у птиц в единую кость — цевку, или плюсну-предплюсну (tarso-metatarsus; рис. 112, 5). Благодаря этому в конечности появляется добавочный рычаг. Подвижное сочленение располагается между двумя рядами костей предплюсны (между костями приросшими к голени, и костями, слившимися с плюсной), поэтому, как и у пресмыкающихся, этот сустав называется интертарзальным (рис. 112, 6).
На дистальном конце цевки образуются хорошо выраженные суставные поверхности для причленения фаланг пальцев (phalanges digitorum; рис. 112, 7). У подавляющего большинства птиц в задней конечности развиваются четыре пальца, из которых I направлен назад, а II, III, IV — вперед.
Мощные длинные кости скелета задних конечностей, появление добавочного рычага (образование tarso-metatarsus), резкий рельеф суставных поверхностей — все это обеспечивает прочность и подвижность задних конечностей.
ЗАКЛЮЧЕНИЕ
Морфологически класс птиц во многих чертах сохраняет большое сходство со своими предками — пресмыкающимися. На это указывает целый ряд признаков: почти полное отсутствие кожных желез, то, что дистальные отделы задних конечностей покрыты роговыми чешуями, роговой покров клюва, диапсидного типа череп (с редуцированной, однако, верхней дугой), имеющий только один затылочный мыщелок, состав костей черепа, то, что имеются шейные ребра и крючковидные отростки грудных ребер, открытый таз (как у некоторых ископаемых динозавров), большое сходство строения мочеполовой системы и периферических отделов кровеносной системы, присутствие клоаки, размножение откладыванием яиц, сходство эмбрионального развития и т. п.
В процессе эволюции птицы обособились от рептилиеобразных предков путем многих, но часто относительно небольших преобразований морфологических структур, обеспечиваюших существенную интенсификацию физиологических функций и тем самым резко повысивших общий уровень организации птиц по сравнению с пресмыкающимися. Полет и высокая общая подвижность птиц связаны с большим расходом энергии, требующим быстрой компенсации.
Поэтому в эволюции птиц большое значение имело становление особенностей организации, связанных не только с обеспечением физической возможности полета, но и с общим повышением интенсивности обмена веществ.
Возможность полета птиц обеспечивается целым рядом прямых приспособлений. К ним относятся: компактность тела, жесткость и неподвижность туловищного отдела осевого скелета и редукция его хвостового отдела, резкое увеличение размеров грудины и образование на ней киля, прочность пояса передних конечностей (в том числе и образование специального амортизирующего устройства — вилочки), строение скелета крыла (включая и особенности суставных поверхностей и особенно резкие изменения в строении кисти), мощность мускулатуры, обеспечивающая движения крыла, развитие перьев — легких и прочных роговых образований, создающих несущие плоскости крыльев и хвоста и обеспечивающих обтекаемость тела в по лете, редукция зубов, облегчающая скелет черепа, общее увеличение объема головного мозга и особенно сильное развитие мозжечка — центра равновесия и координации движений, увеличение массы нервных клеток спинного мозга в области крыльев и т. д.
Клюв с его подвижным скелетным механизмом и дифференцированной мускулатурой стал очень тонко действующим инструментом, способным к сложным движениям. Образование клюва и одновременно увеличение длины и подвижности шеи освободило передние конечности от многих добавочных функций (захват и удерживание добычи, чистка покровов и т. п.) и тем самым способствовало их превращению только в орган полета — крылья. Удлинение шеи и возрастание ее подвижности компенсировало неподвижность туловища и дало возможность, изменяя положение головы, в каких-то пределах менять положение центра тяжести тела, что важно при изменении режима полета.
Преобразование передних конечностей в орган полета — крылья сопровождалось укреплением тазового пояса и скелета задних конечностей, а также возрастанием массы их мышц, обеспечивающих возможность «двуногого» движения на земле, по ветвям деревьев и т. п.
Полет требует затраты большого количества энергии. Например у голубя, летящего со скоростью 70 км/ч, расход энергии в 27 раз больше, чем в покое. Резкое возрастание энергетических возможностей обеспечивается изменениями многих систем органов.
Легкие птиц относительно малы по объему, но велика их губчатость (и тем самым большая поверхность окисления). Сложная система воздушных мешков, связанная с легкими, занимает до 20% объема тела — она резко увеличивает объем вдыхаемого воздуха и дает возможность насыщать кровь кислородом и во время вдоха, и во время выдоха. Чем интенсивнее полет, тем чаще дыхание1, что обеспечивает более интенсивное насыщение крови кислородом и удаление из нее углекислоты при ускорении движения. Одновременно воздушные мешки осуществляют интенсивную теплоотдачу во время полета, предохраняя организм летящей птицы от перегрева.
Полное разделение большого и малого кругов кровообращения резко увеличивает возможность насыщения тканей кислородом. Сердце у птиц относительно очень велико — составляет 0,8—2,5% от веса тела (у рептилий 0,2—0,3%; у млекопитающих 0,2—1,5%). Относительно очень велика у птиц и частота сердцебиений: сердце крупных птиц сокращается 140—200 раз в 1 мин, мелких — до 500—600. В полете частота сердцебиений резко возрастает: так, у голубя в покое около 170 сердцебиений в 1 мин, а в полете 350—600; у мелких птиц частота пульса в полете до 1000 и более. Большой объем сердца и высокая частота сердцебиений создают быструю циркуляцию крови по организму, обеспечивая интенсивное насыщение тканей и органов питательными веществами и кислородом и освобождение их от углекислоты и продуктов распада.
Дифференцировка желудка на два отдела — железистый и мускульный — обеспечивает механическую переработку пищи (в мускульном желудке может развиваться давление до 20—30 кг на 1 см2), что значительно увеличивает интенсивность ее переваривания. Этому же помогает свойственная ряду видов птиц способность отбрасывания погадок, т. е. удаления непереваренных остатков пищи (хитин, шерсть, кости и т. п.) из желудка через пищевод.
Обеспечивает интенсивное пищеварение и высокая активность пищеварительных ферментов. Поэтому скорость переваривания пищи у птиц заметно выше, чем у пресмыкающихся и млекопитающих. Так, воробей переваривает гусеницу за 15 мин, жука — за 1 ч, зерно — за 3—4 ч (считая от времени принятия пищи до вывода ее непереваренных остатков). Интенсивное пищеварение позволяет перерабатывать большое количество пищи, обеспечивая высокие энергетические потребности организма. Особенности пищеварительной системы и органов захвата пищи наряду с большой подвижностью, высоким уровнем высшей нервной деятельности и совершенством органов чувств способствовали образованию широких пищевых связей и создали возможность использования очень большого числа кормовых объектов.
Интенсивность пищеварения, кровообращения и дыхания обеспечивает высокий уровень обмена веществ, что наряду с образованием высоко эффективного теплоизолирующего покрова из пуха и перьев приводит к теплокровности птиц (гомойотермия), т. е. к их способности поддерживать температуру тела на постоянном уровне независимо от изменения температуры окружающей среды. Косвенным показателем высокого уровня обмена веществ птиц может служить температура их тела, которая у крупных видов равна 38—40°С, а у мелких видов достигает 41—43°С.
Значительное увеличение объема головного мозга и возрастание числа нервных клеток в нем, по сравнению с рептилиями, определяют — более высокий уровень высшей нервной деятельности птиц. Это проявляется и в очень сложных системах врожденных рефлексов, определяющих поведение каждой особи, и в значительном возрастании роли индивидуального опыта, т. е. в выработке условных рефлексов.
Плодовитость птиц меньше, чем у пресмыкающихся. Увеличение размеров яиц (откладка которых обеспечивается открытым тазом) увеличивает размеры вылупившегося детеныша. Сопутствующие размножению сложные биологические явления (постройка гнезда, насиживание, выкармливание, обогрев и охрана птенцов и т. п.) обеспечивают ускорение эмбрионального и постэмбрионального развития и повышенное выживание потомства.
Совершенство органов движения (большая подвижность), гомойотермия, сложность высшей нервной деятельности и совершенство органов чувств (особенно зрения и слуха) позволили птицам широко расселиться по всему земному шару. Практически нет на земле ни одного района, включая высокие широты Арктики и Антарктики, где не встречались бы те или иные виды птиц.
Более высокий уровень организации птиц, по сравнению с пресмыкающимися, находит свое отчетливое выражение не только в значительно более широком их распространении по всему земному шару, но и в более сложных и разнообразных взаимоотношениях со средой, с условиями жизни. Так, при сезонных неблагоприятных изменениях условий жизни (наступление зимы) пресмыкающиеся и земноводные прекращают свою активность, впадают в состояние оцепенения. У птиц в целом реакции при наступлении неблагоприятных сезонов года многообразнее и шире. Часть птиц совершает миграции (протяженностью до 5—10 тыс. км и более), улетая на зимовки в благоприятные по погодным и кормовым условиям районы. Однако значительное число видов остается в умеренных и северных широтах в течение всего года. Переживать суровые условия зимы им помогают довольно разнообразные приспособления.
Для многих видов свойственна резко выраженная сезонная смена кормов: переход зимой на питание менее калорийными, но массовыми кормами (например, тетерев летом питается насекомыми, ягодами, зелеными частями растений, зимой — преимущественно почками и сережками некоторых пород деревьев). Неперелетные (оседлые) птицы совершают небольшие перемещения, концентрируясь в участках с большим количеством пищи (глухарь в сосновых лесах, где питается хвоей сосны; синицы, вороны — в населенных пунктах и т. п.). Некоторые виды осенью запасают пищу, пряча ее в укромных местах (кедровки, сойки, поползни — орехи лещины, «орешки» кедра, желуди; синицы — семена деревьев и кустарников). Оседлые птицы во время осенней линьки получают перьевой покров с лучшими теплоизолирующими свойствами, чем летний (большее развитие пуховой части пера, большее число перьев и пуха на единицу поверхности тела и т. п.); у некоторых видов зимний наряд резко отличается по окраске, помогая маскировке (белая куропатка).
У части видов отчетливо выражены и приспособительные особенности поведения: ночевки в укрытиях (тетеревинные и некоторые другие птицы — в снегу; мелкие воробьиные — в дуплах, щелях за корой и т. п.). При этом птицы некоторых видов в сильные морозы ночуют группами, взаимно обогревая в убежище друг друга (синицы, воробьи и др.).
Несмотря на резкое повышение уровня жизнедеятельности и широкие экологические возможности, открываемые гомойотермией, отчетливая специализация птиц к полету в известной мере ограничивает круг экологических приспособлений этих животных. Отсутствие столь узкой специализации у млекопитающих создало предпосылки к возникновению в процессе эволюции этого класса значительно большего разнообразия жизненных форм и более многообразных взаимоотношений со средой.
Дополнительная литература
Гладков Н. А., Дементьев Г. П., Птушенко Е. С., Судиловская А. М. Определитель птиц СССР. М., 1964.
Гуртовой Н. Н., Матвеев Б. С, Дзержинский Ф. Я. Практическая зоотомия позвоночных. Птицы, млекопитающие. М., 1980.
Дементьев Г. П. Птицы. Руководство по зоологии, т. 6. М., 1940.
Дементьев Г. П. Птицы нашей страны. М., 1962.
Иванов А. И., Штегман Б. К. Краткий определитель птиц СССР. Л., 1964.
Карташев Н. Н. Систематика птиц. М., 1974.
Михеев А. В. Биология птиц. М., 1960.
Шмальгаузен И. И. Основы сравнительной анатомии. М., 1947.
Шульпин Л. М. Орнитология. Л., 1940.
Назад Оглавление
Оглавление Далее
Далее
База данных: Каталог библиотеки СФУ (ББК 28.6 Р 851)
Библиографическое описание: Руководство по зоологии / Науч.-исслед. ин-т зоологии Моск. ордена Ленина гос. ун-та им. М. В. Ломоносова.; под ред. Б. С. Матвеева. — Москва ; Ленинград : Издательство Академии наук СССР. — Текст : непосредственный. Т. 6 : Позвоночные. Птицы / сост. Г. П. Дементьев. — 1940. — 856 с. : ил., портр. — Библиогр. в конце разд. — Предм. указ.: с. 802-844. — Имен. указ.: с. 845-856. — (в пер.) : 2.90 р.
Год издания: 1940
Издательство: Издательство Академии наук СССР, Москва ; Ленинград
Количество страниц: 856
Количество экземпляров:
- Абонемент научной литературы (пр. Свободный, 79, 4 этаж, холл): свободно 1 из 1 экземпляров
Ключевые слова: зоология позвоночных, орнитология, классификация отрядов птиц, НЕРВНАЯ СИСТЕМА птиц, нервная деятельность, органы чувств, пищеварительная система, дыхательная система, кровеносная система, географическое распространение, палеоорнитология, эволюция птиц
Рубрики: Зоология / Птицы
Классификационные коды: ББК 28.6, ГРНТИ 34.33
Идентификаторы: полочный индекс ББК 28.6 Р 851, шифр 28/Р 851-735985
Птицы
Расширение среды для осуществления передвижения, способность к полету в
воздухе, присущая представителям более высокоорганизованных, по сравнению с
рептилиями, организмов — именно птиц — связана с
возникновением у них ряда приспособительных прогрессивных морфолого-
анатомических признаков и с появлением более высокого уровня отражения,
психики этих организмов. Птиц следует рассматривать как прогрессивную ветвь
животных, происшедших от пресмыкающихся, приспособившихся к полету в
воздухе.
Наличие передних конечностей, крыльев с летательной поверхностью из перьев,
дает возможность передвижения в воздушном пространстве. Разнообразно
строение задних конечностей птиц — ног, приспособленных к
передвижению в разных средах обитания: в лесах, в открытых пространствах
(луга, степи и пустыни), в водной среде (озера и болота). Челюсти видоизменены
в роговой клюв, имеющий весьма разнообразное строение у разных отрядов птиц
и превосходно приспособленный к схватыванию, удерживанию, убиванию
различного рода живой и захватыванию растительной пищи (ягод, семян
растений). Все это отразилось на поведении птиц, связанном с основной формой
их жизнедеятельности, способами добывания весьма разнообразной пищи.
Естественно, что широта распространения птиц по всему земному шару,
появившиеся широкие возможности соприкосновения и связи птиц с
разнообразнейшими предметами, встречаемыми в весьма различных по характеру
средах их обитания, и обусловили большое развитие их нервной системы и
органов чувств, их ориентировочной деятельности.
У птиц весьма велики большие полушария переднего мозга, а в них —
полосатые тела (Табл. 2.1; 5
); в связи с острым зрением птиц особое развитие получают
зрительные доли их среднего мозга и перекрещивающиеся зрительные нервы.
Перерезка зрительных нервов у голубя с одной стороны вызывает слепоту голубя
на глаз другой стороны. Птица с удаленными полушариями неспособна узнавать
знакомые предметы (например, птица-самка перестает узнавать своих птенцов).
При сравнительно большом объеме полушарий переднего, мозга для птиц
характерна малая величина обонятельных долей (и обусловленное этим слабое
развитие обоняния), большее развитие мозжечка, помогающего координации их
движений и сохранению равновесия птиц при полетах.
Необычайное развитие получили большие по объему глаза птиц, способные к
аккомодации, и не только к монокулярному, но у некоторых птиц и к
бинокулярному зрению. Острота зрения некоторых хищных птиц превосходит
таковую человека. Хищная птица — сокол сапсан (
Falco peregrinus) видит на земле небольших птичек величиной с
горлиц с расстояния свыше 1077 м[84].
Экспериментально доказано, что птицы способны различать цвета так же хорошо,
как человек, но оттенки цвета они видят хуже, чем человек. Они различают
формы предметов (круг, треугольник, прямоугольник), рисунки (горизонтальные
полосы), величины, изображения (в виде разного количества пятен), причем по
степени различения они лучше всего дифференцируют величину, хуже степень
освещенности и цвет и еще хуже форму предметов. Доказано, что птицы
способны различать направление движения предметов.
Столь высоко развитое зрение имеет для птиц огромное значение в связи с их
ориентацией в беспредельном воздушном пространстве при полетах, при
опознавании ими вех на пути возвращения к гнездам, при обнаружении и ловле
живой, тоже летающей добычи, при сезонных перелетах птиц за тысячи
километров от мест их обитания, в связи с наступлением зимы и
неблагоприятными условиями добывания корма.
При испытании слухового анализатора птиц (голубей) оказалось, что они
различали свист, звук рожка, звонок, стук метронома; они дифференцировали
силу, высоту и тембр звуков. Верхняя граница слышимости птицами звуков
лежала между 11 000—12 000 колебаний в секунду. Птицы
дифференцировали высокие тона хуже, чем низкие. В пределах третьей и
четвертой октав голуби были способны различать полутона, в шестой октаве они
различали лишь терции[85].
Слух птиц по отношению к биологически значимым раздражителям значительно
более развит, чем у рептилий; птицы реагировали на еле слышимые человеком
шумы. Тонкость слуха птиц подтверждается их способностью к пению,
тончайшему модулированию своего голоса, различающемуся у разных видов
птиц по высоте, силе звука и ритму, и замечательной способности птиц к
подражанию не только пению и звукам птиц других видов, но и голосам других
животных и даже голосу человека.
Птица-пересмешник может подражать звукам дятла, издаваемым им ударами
клюва по дерезу, мяуканью кошки, воркованью голубя, блеянию ягненка,
карканью ворона. Некоторые вороновые птицы (во́роны,
воро́ны, галки), а также скворцы и особенно попугаи превосходно
имитируют звуки вздоха, кашля, чмоканья губами, щелканья пальцами, свиста и
пения и даже членораздельной речи человека, точно воспроизводя интонацию и
тембр голоса.
Конечно, такое развитие слуха и голосовых способностей имеет для птиц
большое биологическое значение, например, при обнаружении птицами издали
живой добычи, при взаимных поисках самцов и самок в период размножения, при
уходе за молодью. Известны призывные крики птиц, крики тревоги, угрозы и
другие, связанные с выражением эмоциональных состояний птиц и косвенно
служащие для особей своего вида сигналами к проявлению того или другого
характера поведения (например, при подходе к партнеру для спаривания,
избегании опасности, при отпугивании врагов — т. е. при пассивно- или
активно-оборонительных реакциях птиц).
Все эти звуки, конечно, не являются направленными сигналами и выражают лишь
определенные эмоциональные состояния птиц, но они воспринимаются
сочленами группы как биологически значимые для них раздражители среды;
вызывая приспособительные положительные или отрицательные реакции.
Менее развиты анатомически и менее значимы функционально для птиц обонятельный, вкусовой и осязательный рецепторы.
Принимая во внимание сложные формы приспособительного поведения птиц,
относящиеся к питанию, к самозащите, к брачным играм (токам),
сопровождаемым воспроизведением специальных поз, телодвижений (подобия
«танцев»), постройкой своеобразных шалашей и беседок, например,
у шалашниковых райских птиц (Табл. 2.9), наблюдая
замечательное гнездостроение птиц, мы должны определенно признать, что
аналитико-синтетическая деятельность мозга птиц находится на значительно
более высоком уровне по сравнению с таковой у рептилий.
Действительно, мы замечаем у птиц чрезвычайное расширение диапазона
раздражителей, вызывающих деятельность их зрительных и слуховых рецепторов
и интеграцию их восприятий в сложных комплексах эффекторных реакций птиц.
Этими раздражителями, выделяемыми птицами из внешней среды, являются и
живые, и неживые объекты, находящиеся в самых различных средах обитания в
сложном соотношении с окружающим.
Следует подчеркнуть, что добыча птиц чрезвычайно разнообразна, начиная от
млекопитающих, являющихся нередко жертвой крупных хищных птиц, мелких
птиц, рептилий, амфибий, рыб, насекомых, червей и кончая плодами и семенами
разных растений.
Таблица 2.9. Беседка для токования шалашниковых райских птиц

Ловля хищниками добычи в воздухе, на лету, требует большой и точной
координации зрения и кинестезии. Она нередко начинается, когда хищник
находится еще на далеком расстоянии от местонахождения жертвы. Хищная
птица, обозревая с высоты землю, находит притаившуюся живую добычу и не
выпускает ее из поля своего зрения; при передвижении жертвы птица неотступно
фиксирует ее глазами и координирует свою зрительную и двигательную
деятельность, намечая путь полета и спуска, для овладения добычей.
Высматривание некоторыми птицами добычи с воздуха, подкарауливание ее
появления из воды (у хищных птиц, питающихся рыбой, — например,
чаек), вылавливание добычи в воде путем ныряния под водой также требует
тончайшей зрительной дифференцировки и анализа окружающего.
У птиц, питающихся путем процеживания воды сквозь клюв-цедилку (у уток) или посредством обшаривания щелей под корой деревьев клювом и языком (у дятла), в акте питания принимает участие и осязательная рецепция.
У растительноядных птиц, употребляющих в пищу семена растений,
практический анализ окружающих предметов путем зрительных
дифференцировок направлен на различение съедобных для них растений,
выделение частей этих растений (плодов, семян, ягод), на обработку этих
выделенных частей до момента освоения съедобной части.
Особенно интересно, что этот практический анализ при выделении съедобных
частей у разных видов птиц различен по отношению к одному и тому же виду
растений. Например, плод вишни одни птицы (дрозды) поглощают целиком,
другие (воробьи) ощипывают только мякоть плода, третьи (дубоносы)
вылущивают и поглощают лишь косточку плода, бросая мякоть, как несъедобное.
Эти особенности питания наследуются.
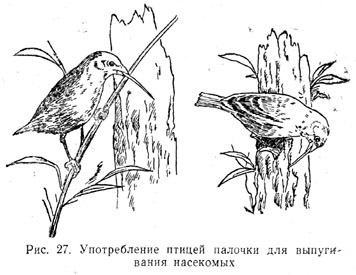
Отметим еще замечательную, носящую прирожденный характер способность
одного вида птиц — именно дятлового древесного вьюрка (Camarhynchus pallidus) — использовать предмет
(кактусовую иглу или веточку) для выпугивания из-под коры дерева насекомых,
которыми эта птица питается. Наблюдатели передают, что птицы обычно
отламывают веточку подходящей длины, отбрасывая слишком короткую или
слишком длинную и заменяя ее соответственной[86]
(Табл. 2.10).
Высокая степень зрительного анализа окружающего у птиц должна проявляться при поисках мест, подходящих для гнездования, в виде готовых убежищ (ямок, норок, дупел, расщелин скал), зарослей кустарника, крон высоких деревьев.
Тонкий, преимущественно зрительный практический анализ сопровождает выбор
птицами подходящего гнездостроительного материала, специфического для
разных видов птиц, — травы, мха, стеблей, прутьев, веток, шерсти, перьев,
паутины, волос, глины, ила и т. д.
Следует подчеркнуть, что птицы, исследованные в условиях неволи методом
естественного эксперимента выдающимся советским орнитологом Промптовым[87]
, обнаружили большую пластичность в выборе гнездостроительного материала,
отступая от стандарта этого выбора, и при наличии искусственного материала
(например, марлевых бинтов), более пригодного для гнездостроения,
использовали его, а не имеющийся поблизости естественный, но менее
пригодный для гнездовития растительный материал.
Таблица 2.10. Употребление птицей палочкидля выпугивания насекомых
Все это определенно указывало, что птицы адаптивна используют свой
индивидуальный опыт, нарушая стереотип видового опыта, инстинкта, т. е.
включают условно-рефлекторную деятельность. На значение индивидуального
опыта и упражнения в процессе конструирования гнезда ярко указывает тот факт,
что молодая птица обычно вьет гнездо хуже старой. Более того, у птиц,
воспитанных изолированно от сообщества, никогда не наблюдавших процесса
стройки, замечается полное смешение гнездостроительных актов, отсутствие их
последовательного и преемственного развития, что указывает на огромную роль
подражания при осуществлении некоторыми птицами гнездостроения (опыты
Промптова).
Сравнительно высокий уровень синтеза раздражителей, получаемых птицами,
обнаруживается в их способности к установлению связей между предметами в
актах конструктивной деятельности, относящейся к постройке замечательных
шалашей для токования и искусных гнезд для выведения птенцов.
Шалашниковые райские птицы сооружают близ ствола дерева своеобразные
круглые высокие арены — платформы из мха, по которым они бегают
кругами во время токования. Другие шалашниковые птицы конструируют
коридоры из прутьев, у входа в которые располагают гладкие камешки, белые
раковины, цветы, заменяя их свежесорванными при увядании (Табл. 2.9).
Как говорит известный советский эколог Д. Н. Кашкаров, у птиц
«эволюция гнездостроения шла в разных систематических группах по
разным путям в связи с особенностями других приспособлений: к средам
обитания, к пище и в связи с особенностями местообитаний»[88].
Вследствие появления у птиц теплокровности, перяного покрова возникло высиживание яиц и выкармливание птенцов, что обеспечивало не только сохранение яиц, но позднее и выращивание птенцов и уход родителей за птенцами.
По развитию птенцов птиц разделяют на две биологические группы — выводковых и птенцовых.
У выводковых птиц (например, куриных, уток и др.) вылупившиеся птенцы в первые же сутки по выходе из яйца покидают гнездо и могут самостоятельно клевать, следуя за матерью.
У птенцовых птиц (голубиных, воробьиных, хищных) птенцы, вылупившиеся из
яйца, довольно беспомощны и долго остаются в гнезде, получая пищу от
родителей. Именно у птенцовых гнезда, устраиваемые родителями, особенно
искусны, так как служат не только для насиживания яиц, но и для выращивания
молодняка в период его выкармливания (Табл. 2.11, Табл. 2.12).
И у птиц легко можно проследить эволюцию инстинкта сохранения потомства и ухода за ним.
Некоторые птицы, гнездящиеся на прибрежных скалах, откладывают свои яйца
просто на землю (гагарка — Alcatorda),
между гальками (крачки, чайки), причем в последнем случае яйца имеют
настолько резко выраженную покровительственную окраску, что их трудно
отличить от окружающих камешков. Другие птицы откладывают яйца в готовых
убежищах: в углублениях земли, в ямках (кулики —
Limicolae), в расщелинах скал, в дуплах (совы — Striges); внутри которых птицы постилают мягкий
материал. Некоторые птицы проявляют большую активность в гнездостроении и
сами вырывают норки в прибрежных пластах песка и земли (береговые ласточки
— Riparia). Гнездостроение, связанное с
долблением твердого субстрата, мы находим у дятлов (
Pici), выдалбливающих себе дупла на стволах деревьев. Далее можно
обнаружить сооружение птицами на земле простейших подстилок под яйца из
мха, травы, листьев (например, у гусей, кур —
Anseres, Galli). Некоторые
группы птиц, также гнездящихся на земле, уже вьют гнездо из трав и стеблей,
например, коньки (Anthus).
Огромное большинство птиц помещают свитые гнезда на ветвях кустарников и
деревьев, то в прикорневой зоне — на тонких ветках дерева, то на высоких
вершинах, где складывают огромные гнезда из толстых сучьев, сверху
застилаемых более мягким материалом (у дневных хищников — орлов, у
врановых).
Для некоторых птиц характерна лепная гнездостроительная деятельность,
связанная с обмазыванием глиной выдолбленного или сплетенного гнезда (у
дроздов — Turdus). Фламинго
воздвигают себе из ила возвышения до 1/2 м
высотой, на верхушке которых делают углубление для помещения яиц и птенцов[89]
(Табл. 2.11). Лепные гнезда, построенные из растений и
склеенные слюной птиц, мы находим у ласточек (Hirunda).
В группе воробьиных птиц (например, у зяблика) можно найти помещаемые на
ветвях деревьев чрезвычайно искусно построенные гнезда, сплетенные из
различных материалов: толстые стенки гнезда сделаны из мха, тонких корней и
прутиков; внутри гнездо выстлано волосом и перьями, а снаружи облицовано
лишайниками и берестой под тон того ствола дерева, на котором гнездо
находится, что хорошо скрывает гнездо на окружающем фоне.
Особенно искусно бывают сделаны гнезда камышевок, имеющие форму глубоких
цилиндрических вместилищ, висящих среди вертикальных стеблей камышей,
гнезда тропических ткачиков, сшитые из сближенных и скрепленных между
собой листьев, и гнезда синицы-ремеза в виде варежки (Табл. 2.12) — из мягкого растительного материала.
Таблица 2.11. Гнездо фламинго

Самка птицы-носорога откладывает яйца в дупла, где самец замуровывает ее глиной на все время высиживания птенцов; он кормит ее, освобождая из гнезда лишь ко времени вылупления птенцов.
Иногда при постройке гнезд сочетается роющая и конструктивная деятельность птиц.
Замечательны гнезда сорных кур (Megapodidae
), зарывающих свои яйца внутрь огромных куч (высотой до 110
см) растительных веществ, где через 45 дней,
под действием развивающегося от гниения тепла, выводятся птенцы. Молодая
птичка разбивает клювом верхний тупой конец яйца, вылупляется и с помощью
крыльев выбирается наружу, кверху. Как только она достигает поверхности
холма, она зевает, отряхивается и бежит прямо в соседние кусты. Но если такого
птенца вторично зарыть в кучу, то он оттуда не может выбраться и погибает.
Таблица 2.12. Гнезда птиц различных видов

Таким образом, конструктивная деятельность птиц достигает большой сложности
в актах гнездостроения, начинаясь от более Примитивной стадии устройства
простой подстилки для яиц из травы, листьев, мха, располагаемых на земле, и до
более сложного сооружения, когда гнездовое вместилище конструируется птицей
путем свивания, сплетения (стеблей, мха, волокон растений, водорослей),
сшивания из листьев, склеивания из грязи, глины, слепления затвердевающей
слюной.
Из этих разнообразнейших материалов птицы устраивают разной сложности и
разной формы гнезда в виде лоточка, корзинки, шара, реторты, варежки, чулка и
т. д. (см. Табл. 2.12). Гнезда некоторых видов птиц облепляются
сверху дополнительным материалом — листочками, растительными
чешуйками, мхом, хорошо скрывающими гнездо на окружающем его фоне.
Анализ гнездостроительной активности птиц А. Н. Промптовым привел его к
заключению, что то, что называют инстинктивным поведением птиц, есть
равнодействующая сложнейшего сочетания прирожденных
безусловнорефлекторных и условнорефлекторных реакций.
«Основная и характернейшая черта этого поведения — направленная
активность — как раз и иллюстрирует это положение, так как она
обусловливается течением и сменой различных биологически-адаптированных
доминант, функционально объединяющих разные „этажи“ нервной
системы и тесно связанных также с гуморальной динамикой организма»[90].
«Целесообразность же инстинктивного поведения, — согласно
Промптову, — имеет глубоко эволюционные корни, так как по сути дела
зависит от функционирования врожденных механизмов, отобранных и
закрепленных длительным процессом естественного отбора. Взаимодействие
этих, филогенетически сложившихся (прирожденных) компонентов поведения с
онтогенетически и индивидуально развивающимися компонентами,
обусловливает степень приспособления каждого вида, определяет формы его
реагирования на меняющиеся условия среды…».
«В таком дарвиновском смысле, понимаем мы, — пишет А. Н.
Промптов, — эволюцию инстинктов, которые, конечно, меняются,
затухают, исчезают совсем или рождаются вновь, в неразрывной связи с анатомо-
морфологической эволюцией вида, текущей в характерной для него и также
эволюционирующей внешней среде»[91].
Но, отмечая высокое развитие конструктивной деятельности птиц, мы должны
подчеркнуть, что эти их способности проявляются при определенных
нормальных условиях жизнедеятельности. Отклонение от нормы вызывает
нецелесообразные действия птиц, свидетельствующие о ситуационной
связанности их восприятий, в общем о стереотипности их сложной
инстинктивной деятельности. Например, если у чаек, гнездящихся на земле среди
прибрежных камней, переложить яйца из ямки, где они лежали, в другое место, то
птицы продолжают насиживать пустую ямку, хотя яйца лежат поодаль, но близ
нее; если в той же гнездовой ямке яйца заменить камнями или другими
округлыми деревянными предметами, птица-мать этого не замечает и продолжает
насиживание[92].
Аналогичные опыты были проведены Б. И. Хотиным и Г. З. Рогинским над
кайрами на птичьих базарах Новой Земли[93].
По сравнению с рептилиями у птиц в проявлении инстинкта охраны потомства
появляются новые, гораздо более длительные и активные связи матери, а иногда
и; обоих родителей с подрастающим поколением. Как уже было отмечено,
птенцовые продолжительно, выкармливают детенышей, приучают их к полету и
ловле живой добычи (у хищных птиц) посредством бросания родителями в
воздух пойманной живой жертвы перед птенцами; таким образом и
предоставляется возможность поимки добычи молодыми птицами. В этом случае
имеет место установление у птиц сложных связей уже не только между особями
своего вида и своим потомством, но и межвидовых отношений с другого вида
животными.
Исследования А. Н. Промптова, произведенные в Колтушской лаборатории (1947
г.), показали, что биологический контакт поколений играет очень большую роль в
жизни птиц, в формировании их видового стереотипа поведения. Птенцы,
находящиеся в непосредственной связи с выкармливающими их родителями, на
основе, «свойственной всем птицам способности к подражанию…
„осваивают“ характерные для вида кормовые стации, виды
кормов…»[94].
И этого типа связи, хотя и осуществляются в основном в русле видовой
инстинктивной деятельности, но, конечно, у каждой особи в процессе ее развития
они образуются на основе индивидуального опыта и довольно сложной
аналитико-синтетической деятельности ее мозга.
В последнее время, работая также в Колтушской лаборатории, Е. В. Лукина[95]
, анализируя взаимодействие врожденных и приобретенных реакций птиц при
выявлении изменяющегося воздействия среды обитания на формирование их
пищедобывательной и гнездостроительной деятельности, пришла к интересным
выводам. Она доказала, что под влиянием человека при перемене условий
содержания птиц в различных ландшафтах можно формировать
пищедобывательные реакции некоторых птиц, изменяя их стереотипный видовой
способ питания на новый. Точно так же возможно заставить птиц изменять место
гнездования и стереотипный выбор определенного гнездового материала.
Означенное, направленное изменение инстинктов птиц можно широко
использовать в практическом отношении, например, при необходимости
переселения птиц в лесозащитные полосы.
В жизни птиц наблюдается интереснейшее явление — это весенние и осенние перелеты, т. е. регулярные массовые их переселения из одной области в другую.
С наступлением весны из жарких стран к нам в среднюю полосу СССР начинают прилетать различные птицы.
«Весенний перелет, — пишет А. Н. Промптов, — открывается
появлением грачей. Затем прилетают полевые жаворонки, скворцы и вслед за
этими первыми переселенцами стремительно набегает волна массового появления
сразу многих видов перелетных птиц»[96].
В. Н. Бостанжогло в своей большой работе о птицах Арало-Каспийских степей (1911) так описывает валовой весенний пролет в устье р. Урала:
«Уже с половины марта и почти весь апрель (ст. ст.) горизонт ни на минуту
не остается свободным. Трудно даже сказать, в какое время дня и ночи птицы
летят меньше, так как стон стоит в воздухе во всякий час. Торопливо несутся
развернутым фронтом стаи всевозможных пород уток, выплывая из-за горизонта
длинными темными линиями и с тревожным свистом проносясь над устьями
реки; тесными клубками валят стайки куликов и ржанок, перегоняя грациозных
кроншнепов и суетливых веретенников; озабоченно пролетают одинокие
хищнику; мерно и величественно плывут цапли. В приморской полосе в это время
птица собирается в невероятном количестве. Подобно вокзалу большой узловой
станции, все полно движения, сутолоки, криков, но нетребовательны пассажиры и
дорого их время. Переведя дух и заморив на скорую руку червяка, стаи
снимаются и летят далее, сменяясь все вновь прибывающими. Только немногие
птицы делают более продолжительные остановки и не торопятся с окончанием
своего воздушного путешествия. Во второй половине апреля в устьях Урала
наступает уже сравнительная тишина, и местные птицы уже давно оберегают
занятые участки…»[97].
С конца лета обычно у птиц начинается новое великое переселение.
«Стаями и в одиночку, днем, ночью, утренними или вечерними зорями,
каждый вид со свойственными ему особенностями, двигаются птицы на зимовки
— снова в те места, откуда они прилетели несколько месяцев назад. Теперь
эта армия переселенцев еще многочисленнее — она сильно пополнилась
молодняком, который впервые в жизни совершает полет»[98].
«
Осенью от нас улетают миллионы птиц. Конечно, многие гибнут по пути
от бесчисленных опасностей, связанных с перелетом. За сотни и тысячи
километров от своих гнездований они летят до тропической Африки, Индии,
Китая и Индокитая или же задерживаются на побережьях Средиземного моря или
у южных берегов Каспия.
В этих местах находятся зимовки большинства птиц, населяющих летом Европу и
север Азии. На зимовках перелетные птицы находят себе подходящую пищу,
удобные условия жизни и держатся там до весны[99].
»
Большинство птиц, пролетая во время весеннего перелета тысячи километров,
возвращается именно в те места, где они вывелись. Если осенний перелет птиц
вызывается сезонными изменениями внешних условий: наступлением холодов и
уменьшением кормов, то весенний прилет с мест зимовок вызывается в
значительной степени внутренним физиологическим состоянием организма,
состоянием, предшествующим периоду размножения птиц, связанного с
определенными экологическими условиями — с поисками традиционной
для вида гнездовой стации, — приводящими птиц в область старого
гнездования, даже прошлогоднего гнезда.
Чем же руководствуются птицы в своих сезонных передвижениях, пролетая огромные пространства между двумя совершенно определенными географическими областями?
В этом отношении имеется много разных предположений. Одни ученые
выдвигают значение физических факторов, вплоть до влияния земного
магнетизма; некоторые ученые (Цион) на основании исследования ориентировки
в пространстве почтовых голубей ссылаются на наличие у птиц специальной
чувствительности к восприятию температурных изменений, интенсивности и
направления ветра, ориентирующих птицу при полете. Ряд исследователей
придает большое значение зрению птиц, их прекрасной зрительной памяти.
Другие авторы, занимавшиеся этим вопросом, подчеркивают связь зрительного и
мускульного запоминания общей трассы перелета, или объединения восприятия
нескольких органов при учете долготно-широтных координат при ориентировке
птиц в пути. Пути перелета птиц являются определенными, складывавшимися
тысячелетиями.
В перелетах птиц имеет громадное значение видовая инстинктивная традиция,
передаваемая от поколения к поколению во время совместных странствований
старых птиц с молодыми. Этот миграционный инстинкт является пережитком
далекого прошлого птиц и благодаря ему древние зимовки, иногда, вследствие
расселения вида, ставшие очень далекими, остаются все еще прочно связанными
с его современным гнездовым ареалом.
Справедливо говорит А. Н. Промптов, что для разработки вопросов миграции
необходим комплексный метод исследования на основе данных зоогеографии,
экологии, физиологии и генетики. Это сложное явление не может быть правильно
понято одним анализом составляющих его компонентов, изучаемых разными
методами этих наук.
Переходя к особенностям индивидуально приобретенного, собственно
условнорефлекторного поведения птиц, как оно обнаруживается в
экспериментальной ситуации[100]
, следует сказать, что самые различные зрительные раздражители могут быть для
птиц сигнальными положительными раздражителями, например, свет, темнота,
цвет, форма, величина фигур, количество пятен, направление движущихся фигур.
В своих экспериментальных исследованиях[101]
зрительных дифференцировок у различных видов попугаев мы имели
убедительные доказательства, что попугаи: ара, лори, какаду легко вырабатывают
условные рефлексы на тот или другой цвет: красный, синий, зеленый,
помещенный среди различных цветовых компонентов (Табл. 2.13).
В Колтушской лаборатории посредством применения «хватательной
методики», в частности вызова пищедобывательного движения птиц (кур,
голубей, ворон, галок, уток) путем дерганья клювом кольца или нажима клювом
на рычаг, удалось выработать у птиц стойкие условные рефлексы и
дифференцировку самых различных зрительных и слуховых раздражителей
— зеленого и белого цвета, звука зуммера, метронома, звонка, —
сочетаемых с подкреплением пищей. Следует отметить, что эти рефлексы
сохранились после 5-месячного перерыва в работе с птицами[102].
При сравнении деятельности различных анализаторов птиц в процессе выработки
условных рефлексов на дифференцировку раздражителей оказалось, что
наилучшее дифференцирование давали зрительный и слуховой анализаторы[103]
. Значительно хуже осуществлялась обонятельная дифференцировка различных
запахов (одеколона, ментола, валерьяны); еще хуже действовал
пространственный анализатор (дифференцировка направления вращения) и
временной (образование условного рефлекса на получасовые промежутки).
У журавлей (Grus collarus) в зоологическом
саду (в Галле) был выработан условный рефлекс приходить на свист к месту
кормежки. Этот навык сохранился вопреки перерыву в подаче свиста в течение
21½ месяца.
В одном совхозе у гусей образовался условный рефлекс — целыми стаями
прибегать с озера по звону колокола, извещавшего рабочих о наступлении часа
обеденного перерыва. Куры выучились находить кратчайший путь в лабиринте,
при прохождении которого надо было приобрести (выбирая один путь из двух) 23
различных условных рефлекса, составляющих сложную ассоциативную связь,
обеспечивающую решение задачи. Но в более сложной обстановке куры,
например, не могли найти обходный путь к пище, брошенной за решеткой, хотя
требовалось только отойти от пищи и обойти решетку (и этот путь был им
известен). Собака, а тем более обезьяна, легко решала подобную задачу (опыты В.
Кёлера)[104].
Много опытов было поставлено с целью проверить способность различных видов
птиц к ориентировке в лабиринте, к освоению и запоминанию ими кратчайшего
пути выхода из лабиринта (опыты М. П. Садовниковой)[105].
На основании этих исследований установлено, что при ориентировке в лабиринте
у снегиря вначале ведущую роль играет зрение, позднее, после освоения с
лабиринтом, кинестезия. Изменение положения того же лабиринта — его
поворот на 90°, 180°, 270° дезориентирует птицу, и она
путается в прохождении; только в результате новых опытов она опять находит
правильный и кратчайший путь выхода.
Таблица 2.13. Выбор попугаем какаду цветной пластинки

Испытанные по методу лабиринта голуби после прохождения одного лабиринта легче научились находить путь во втором лабиринте, разнящемся по конструкции от первого (опыты Роузе).
Голубей приучали открывать дверцу ящика с кормом, а воробьи были
вытренированы открывать дверцу клетки с кормом путем потягивания ее за
ниточку. Птицу-зеленушку приучили выбирать пищу из ящика, (находящегося
среди четырех), причем она находила этот ящик и при перемене его
местонахождения (опыты Бойтендайка).
Вороны оказались способными к образованию условного рефлекса на выбор
определенного в ряду предмета. Так, из девяти открытых дверей они выбирали
заданные: крайнюю правую или левую, но они не могли освоить выбор второй с
краю левой двери[106].
Академик Бериташвили исследовал способность голубей к восприятию внешнего
вида пищи, состоящей из большего (шести) или меньшего (трех) количества
зерен. Опыты показали, что сначала голуби неизменно начинали клевать кучу,
содержащую большее количество (шесть зерен); позднее они направлялись к
кучке, содержащей меньшее количество (три зерна).
Когда же голубей движением руки стали отгонять от большей кучи, то через 250—260 опытов у них выработали условный рефлекс на клевание из меньшей кучки.
После 1001—1413 кормлений из кучки с тремя зернами голубь тянулся к
этой кормушке, независимо от того, какое количество зерен находилось в
соседней. Этот условный рефлекс закрепился настолько прочно, что вне клетки
голубь клевал определенную кучку, состоящую из трех зерен; к другим кучкам он
или не подходил, или подходил с большим промедлением[107].
Немецкий ученый О. Кёлер, исследовавший способность птиц к различению количества предметов, пришел к заключению, что птицы могут различать количество, определяя его «на глаз».
В его опытах голуби отличали пять зернышек от шести, когда обе группы
располагались рядом. Более того, они приучились выклевывать из кучки только
пять зерен, не трогая оставшиеся зерна. Волнистые попугаи выучивались
различать количество шесть, и это количество было пределом их различения, а
ворон, серый попугай и амазонский попугай могли различать до семи единиц.
Ворон, с которым О. Кёлер работал по нашей методике «выбора на
образец», научился на показываемый ему образец-кружок с
3—4—5—6—7 черными точками открывать ту
кормушку, на которой было изображено соответствующее количество точек[108].
Экспериментальное изучение способности к образованию условных рефлексов у птиц, в частности, у воробьев, дает возможности выявить некоторые их особенности при решении разного рода задач.
Такими особенностями являются: использование птицами своего
индивидуального опыта, подражание одних индивидов опыту других при
решении задач, наличие пробных, поисковых опытов, предваряющих правильное
решение задачи, большая настойчивость в преодолении препятствий, способность
переучиваться, приметливость к появлению новых элементов на фоне прежних,
адаптация своих действий к меняющимся условиям, наличие хорошей памяти[109].
Поставленные в сравнительно-физиологическом плане опыты по изучению
условных рефлексов у представителей позвоночных разных классов: рыб, черепах
и птиц (галки) — при анализе подвижности корковых нервных процессов
выявили возможность переделки у этих животных сигнального значения
раздражителей, (именно зеленого и белого цвета), но повторная переделка лучше
удалась у птиц, меньше у черепах и совсем не удалась у рыб[110]
. Это происходило, по мнению автора, вследствие того, что птицы обладают
большой тренируемостью подвижности нервных процессов по сравнению с
черепахами и рыбами. У птиц удалось также выработать условные рефлексы
второго порядка.
Баяндуров, сравнивая скорость образования условных рефлексов и их прочность
у представителей пяти классов животных: рыб, амфибий, рептилий (черепах), а
также птиц и млекопитающих (собак), подчеркивает, что чем выше в
зоологической лестнице стоит животное, тем сильнее в его центральной нервной
системе выражены тормозные процессы.
У собак они довольно значительны, гораздо интенсивнее, чем у птиц, у птиц
значительнее, чем у рыб, амфибий или рептилий. И, наоборот, чем ниже на
зоологической лестнице стоит животное, тем в его центральной нервной системе
хуже протекают процессы внутреннего торможения условных рефлексов, а
отсюда худшая дифференцировочная способность.
Согласно мнению Баяндурова, по способностям к образованию условных
рефлексов птицы занимают среднее место между рыбами и млекопитающими и
далеко отходят от амфибий и рептилий[111].
Обзор способности птиц к установлению условнорефлекторных связей в
экспериментальной ситуации, выделение ими индифферентных раздражителей,
сочетаемых с безусловным раздражителем, оценка точности дифференцировок
раздражителей, адресованных к разным анализаторам, выявление прочности
сохранения образованных навыков, их переделки при изменении значения
раздражителей в новых ситуациях позволяет нам сделать заключение о характере
аналитико-синтетических процессов у птиц.
Совокупность имеющихся экспериментальных данных обнаруживает сильно развитую способность птиц к аналитическому выделению самых различных по качеству раздражителей.
Обнаруживается способность птиц не только к выделению, но и тонкой
дифференцировке многообразных раздражителей (зрительных, звуковых, в
меньшей степени обонятельных, пространственных, временных) на фоне
окружающей обстановки, что связано с участием тормозных процессов, которые
у птиц явно более выражены, чем у рыб, амфибий и рептилий.
Совершенно несомненна довольно высокая способность птиц не только к
синтезированию индифферентных раздражителей с биологически значимыми, но
связывание их сенсорных дифференцировок с разнообразнейшей двигательной
активностью птиц — движением клюва, ног, с актами дерганья, нажима,
схватывания клювом, взятия и подачи клювом, с подбеганием к определенному
раздражителю, прохождением лабиринта, подлетом и присаживанием на жердь и
т. д.
Установленные синтетические связи птиц отличались большой прочностью.
Выучка сохранялась у птиц при перерыве в работе от 5 до 21 месяца, в
противоположность тому, что наблюдалось у амфибий (лягушек), у которых
выработанные условные рефлексы приходилось образовывать заново в каждый
последующий день экспериментов, и у рептилий (черепах), у которых угасание
условного рефлекса шло медленнее, чем у амфибий, но скорее, чем у птиц.
Вопреки относительно большой устойчивости, установленные временные связи
птиц отличались пластичностью и могли быть видоизменены, переделаны, а у
испытанных в лаборатории рептилий (черепах), и в еще большей степени у рыб и
амфибий такая переделка давалась с трудом.
С другой стороны, следует подчеркнуть, что в случае установления более
сложных синтетических связей, последние, возникали у птиц в результате более
или менее длительного поискового пути (проб и ошибок), предваряющего
образование замыкательного процесса в мозгу птиц, обеспечивающего
установление условной связи.
Эти установленные временные связи были действенны преимущественно в
определенной ситуации, в которой они были получены, перенос их в измененные
условия, сравнительно мало разнящиеся от прежних, содействовал более скорому
образованию новых связей на базе существующих, например, при осложнении
пути при ориентировке птицы в лабиринте.
Но при резкой перемене условий, например, повороте лабиринта на
90°—270°, требующей более радикальной перестройки
образовавшихся двигательных навыков, птицы не могли сразу решить задачу.
Птица снова нащупывала измененный путь, пробега, снова устанавливала связи
для образования соответственного нового условного рефлекса. Прочно
фиксированные у птиц, сравнительно мало пластичные условные связи в более
сложных случаях препятствовали образованию новых условных связей. Этим,
объясняется трудность доставания птицами корма, обнаруженная, например, у
кур, при необходимости использования ими обходных путей. Как было
упомянуто, курица, привыкшая брать видимый корм, непосредственно перед ней
расположенный, настойчиво рвалась через сетку к этому корму, вопреки
имеющемуся у нее (на основании прежнего опыта) знанию других путей выхода к
корму; она явно уступала в отношении пластичности условных связей
млекопитающим животным (собаке и обезьяне)[112].
Сравнивая уровень отражения у птиц с таковым ранее рассмотренных позвоночных животных (рыб, амфибий, рептилий), мы находим у них несомненное повышение и усложнение форм отражения.
Появление коры мозга и совершенствование высокоорганизованных органов
чувств — зрения и слуха — определяет значительный прогресс
анализа, особенно зрительного, как в отношении его необычайного обострения,
так и возникновения цветного и предметного зрения. Аналогичное утончение и
расширение диапазона рецепции имеется у птиц и в области слуха. В
соответствии с этим наблюдается значительное расширение ориентировочной
деятельности птиц. Аналитико-синтетическая деятельность мозга птиц
приобретает более сложные формы вследствие возрастания многообразия связей
организма со средой.
Птицы оказались способными к уточненному анализу отдельных свойств
изолированных раздражителей окружающей среды как зрительных (света, тьмы,
яркости, цвета, формы, величины, рисунка, движения и направления движения
предметов), так и слуховых (стука, свиста, шума и высоты, силы, тембра, ритма
звука).
Птицы оказались способными к синтезу получаемых раздражений в целостное
восприятие при зрительном различении предметов, при подражательной
репродукции тех же видов звуков, при воспроизведении ими голосов различных
птиц и других животных и даже членораздельной человеческой речи.
Сложная аналитико-синтетическая деятельность птиц проявляется в актах
питания, самосохранения, размножения, сохранения потомства, включающих
участие огромного количества безусловных сигнальных раздражителей,
вызывающих биологически целесообразные реакции птиц.
Напомним сложно осуществляемые акты питания, связанные, например, с
высматриванием добычи, выбором съедобных плодов, акты конструирования
(шалашей, беседок, платформ для толкования, гнезд для выведения птенцов),
когда птица избирательно выделяет материал из сложного комплекса
окружающей среды, когда она многообразными приемами осуществляет связи
между самыми различными предметами, оформляя самые различные типы гнезд.
В отношении деятельности гнездостроения следует отметить, что наравне с
натуральными, сигнальными раздражителями, строго определенными у разных
видов птиц, оказываются действительными в силу генерализации раздражителей
и другие, новые сигнальные стимулы, более или менее сходные с обычными. Так,
например, птицы могли использовать для сплетения гнезд вместо стеблей
растений случайно оказавшиеся поблизости предметы человеческого обихода
— бинты — и даже предпочитали эти материалы естественным.
Подобное изменение поведения указывает на возможность нарушения птицами
видового динамического стереотипа при изменении внешних условий и на
способность их к широкому использованию своего индивидуального опыта.
Прогрессивные особенности форм отражения птиц ярко сказываются в их
способности к выработке навыков, т. е. условных рефлексов на самые различные
индифферентные раздражители, сочетаемые с безусловными и ставшие
сигнальными. Многообразие условных сигнальных раздражителей (зрительных,
слуховых, обонятельных) свидетельствовало о том, что птицы были в состоянии
осуществлять более дробный и углубленный по сравнению с рептилиями анализ
новых свойств и качеств среды и более сложный новый синтез раздражителей на
основе накопления своего индивидуального опыта и приспособительного его
использования. Выработка у птиц условных рефлексов, связанная с различением
ими многообразных зрительных и слуховых сигнальных признаков, так же как и
при нахождении ими пути в лабиринте, указывает, что аналитико-синтетические
процессы птиц состоят не только в дифференцировке ими отдельных свойств
предметов, но и в целостном восприятии ими совокупности этих свойств, в
восприятии комплексных раздражителей, и их соотношений в длительном,
прочном их сохранении.
У птиц, по сравнению с рептилиями, сильнее выражены и тормозные процессы
мозга и подвижность нервных процессов при переделке сигнального значения
раздражителей. Синтез птицами индифферентных раздражителей в сложном
интегрировании их в самых различных условнорефлекторных связях
свидетельствует и об усложнении динамики нервных процессов птиц и более
сложном взаимоотношении и взаимодействии механизмов, являющихся
физиологической основой весьма углубившегося и уточнившегося отражения
окружающего мира.
Но все же следует подчеркнуть сравнительно малую подвижность высшей
нервной деятельности птиц, недостаточную пластичность их поведения при
резкой перемене условий, отражающуюся в трудности переделки прежних
условных связей и образования новых.
Карташев Н.Н., Соколов В.Е., Шилов И.А.
Практикум по зоологии позвоночных
М., Высшая школа. 1981 г.
ТЕМА 17. СКЕЛЕТ ПТИЦЫ
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ ОБЪЕКТА
Подтип Позвоночные, Vertebrata
Класс Птицы, Aves
Отряд Воробьинообразные, Passeriformes
Представитель — Серая ворона, Corvus corone L.
МАТЕРИАЛ И ОБОРУДОВАНИЕ
Для занятий важно иметь хотя бы один смонтированный скелет птицы. На одного-двух студентов необходимы:
1. Разборный скелет вороны — 1 комплект.
2. Препарировальные иглы — 2.
ЗАДАНИЕ
Уяснить особенности строения скелета птиц. Сделать следующие рисунки:
1. Череп вороны сбоку.
2. Череп вороны снизу.
3. Скелет туловища сбоку (включая позвоночный столб, пояс передних конечностей, грудину, ребра, тазовый пояс).
4. Тазовый пояс и сложный крестец снизу.
5. Скелет крыла.
6. Скелет задней конечности.
7. Первый, второй и один из задних шейных позвонков (вид сбоку).
ОПИСАНИЕ СКЕЛЕТА
В скелете птиц отчетливо выражены черты, связанные с приспособлением к полету. Плоские кости обычно очень тонкие, губчатые. В больших трубчатых костях хорошо развиты полости, заполненные или костным мозгом, или воздухом. Все это обеспечивает повышенную прочность скелета и заметно облегчает его. Нужно, однако, иметь в виду, что общая масса скелета птиц составляет 8—18% от массы тела, т. е. примерно столько же, сколько и у млекопитающих (6—14% веса тела), хотя у последних кости толще, а воздушные полости в них отсутствуют. Это объясняется тем, что у птиц значительно больше относительная длина скелетных элементов конечностей и некоторых других костей.
Скелет птиц подразделяется на осевой скелет, грудную клетку (грудина и ребра), череп, скелет конечностей и их поясов.
Осевой скелет. Осевой скелет, или позвоночный столб (column. vertebralis), как и у пресмыкающихся, разделяется на пять отделов шейный, грудной, поясничный, крестцовый и хвостовой.
Шейный отдел очень подвижен (птицы легко поворачивают голов на 180°, некоторые даже на 270°). Число шейных позвонков варьирует в широких пределах: от 11 (попугаи) до 23—25 (лебеди, утки); у вороны, голубя 14 шейных позвонков.
Рис. 105. Шейные позвонки вороны
А — атлант; Б — эпистрофей; В — средний шейный позвонок:
1 — сочленовная ямка для затылочного мыщелка, 2 —канал для спинного мозга, 3 — тело позвонка, 4 — зубовидный отросток, 5 — верхняя дуга, 6 — остистый отросток, 7 — передние сочленовные отростки, 8 — задние сочленовные отростки, 9 — поперечный отросток, 10 — шейное ребро, 11 — позвоночное отверстие
Первый шейный позвонок — атлас, или атлант (atlas; рис. 105, А) имеет форму кольца. На нижней части его передней поверхности есть глубокая сочленовная ямка, в которую входит затылочный мыщелок черепа; на нижней задней поверхности — сочленовная плоскость для соединения с телом второго шейного позвонка. Просвет полости кольца поперечной сухожильной, обычно окостеневающей связкой разделен на две части: через верхнюю проходит спинной мозг, а в нижней помещается зубовидный отросток второго позвонка.
Второй шейный позвонок — эпистрофей (epistrophaeus; рис. 105, Б) имеет хорошо развитое тело с направленным вперед зубовидным отростком (рис. 105, 4). По происхождению зубовидный отросток, видимо, — тело первого позвонка, обособившееся от него и сросшееся с телом второго позвонка. Сбоку расположены поперечные отростки; к ним крепятся шейные мышцы, соединяющие второй позвонок с затылочным отделом черепа и с другими шейными позвонками. Над телом позвонка расположена хорошо развитая верхняя дуга (рис. 105, 5), через просвет которой проходит спинной мозг.
Остальные шейные позвонки имеют гетероцельное строение: довольно длинное тело каждого позвонка (corpus vertebrae; рис. 105, 3) на передней и задней стороне имеет седловидные сочленовные поверхности. Такой характер сочленения обеспечивает значительную подвижность позвонков друг относительно друга, как в вертикальной, Так и в горизонтальной плоскостях. Это обеспечивает наряду со сложной системой дифференцированных шейных мышц большую подвижность шеи, что очень важно для птиц (компенсация неподвижного скелета туловища, возможность менять положение центра тяжести в полете, сгибая и выпрямляя шею, возможность сложных движений головы при чистке оперения, постройке гнезда и т. п.).
Над телом позвонка расположена верхняя дуга (arcus neuralis; рис. 105, 5), заканчивающаяся остистым отростком (processus spinosus; рис. 105, 6); в канале дуги проходит спинной мозг. От передней и задней поверхности дуги отходят парные сочленовные отростки (рис. 105, 7, 8), соединяющиеся с такими же отростками соседних позвонков. Такой тип сочленения обеспечивает прочность соединения позвонков друг с другом при сохранении значительной подвижности. От боковой поверхности тел позвонков отходят короткие поперечные отростки (processus transversus; рис. 105, 5). Птицы, как и пресмыкающиеся, имеют шейные ребра (рис. 105, 10); они рудиментарны и только последние одно-два шейных ребра достаточно длинны и подвижны (рис. 106, 5), но не доходят до грудины. Шейное ребро прирастает своей головкой к телу позвонка, а бугорком — к поперечному отростку. Таким путем образуются позвоночные отверстия (foramen transversarium; рис. 105, 1), создающие с обеих сторон шеи канал, по которому проходят позвоночная артерия и шейный симпатический нерв. Эта костная защита артерии и нерва особенно важна при длинной, очень подвижной шее.
Рис. 106. Скелет туловища вороны сбоку:
1 — последние шейные позвонки, 2 — сросшиеся грудные позвонки (спинная кость), 3 — шейные ребра, 4 — грудные ребра, 5 — крючковидный отросток, 6 — тело грудины, 7 — киль, грудины, 8 — сложный крестец, 9 — подвижные хвостовые позвонки, 10 — пигостиль, 11 — коракоид, 12 — вилочка, 13 — лопатка, 14 — суставная впадина для головки плеча, 15 — подвздошная кость,
16 — седалищная кость, 17 — лобковая кость, 18 — вертлужная впадина для сочленения с головкой бедра
Грудной отдел у вороны представлен шестью позвонками (у других видов птиц их число варьирует от 3 до 10), которые, как и у большинства птиц, срастаются друг с другом, образуя спинную кость (dorsale; рис. 106, 2).
К грудным позвонкам подвижно причленяются грудные ребра (costa; рис. 106, 4); число их пар равно числу грудных позвонков. Это плоские, слегка изогнутые костные пластинки, состоящие из двух подвижно соединенных друг с другом отделов — спинного и брюшного. Верхний конец спинного отдела ребра имеет два сочленовных отростка: бугорок, причленяющийся к поперечному отростку, и головку, сочленяющуюся с телом позвонка. Нижний конец брюшного отдела ребра подвижно сочленяется с телом грудины (sternum рис. 106, 6). Подвижное соединение ребер с грудиной и позвоночным столбом вместе с подвижным сочленением спинного и брюшного отделов ребра обеспечивает возможность значительных изменений объема полости тела путем опускания грудины вниз и поднимания ее кверху. Эта подвижность возможна благодаря сильно развитой реберной мускулатуре. Жесткость грудной клетки усиливается костными образованиями — крючковидными отростками (processus uncinatus; рис. 106, 5) ребер, укрепляющимися на спинном отделе и налегающими на последующее ребро.
Грудина, или грудная кость (sternum; рис. 106, 6) — широкая и длинная, вогнутая изнутри костная пластинка, несущая по средней линии высокий костный гребень — киль грудины (crista sterni; рис. 106, 7). Резкое увеличение поверхности грудины, обусловленное как ее разрастанием, так и образованием киля, — прямое приспособление к полету; оно обеспечивает возможность прикрепления мощных мышц, приводящих в движение крыло. На переднем конце грудины по бокам киля расположены большие сочленовные площадки, обеспечивающие прочное прикрепление к телу грудины коракоидов (рис. 106, 11). На боковых краях грудины размещаются мелкие углубления, которые служат для сочленения с нижними концами брюшных отделов ребер.
Рис. 107. Сложный крестец вороны (вид снизу):
1 — поясничные позвонки, 2 — крестцовые позвонки, 3 — хвостовые позвонки,
4 — подвздошная кость, 5 — седалищная кость, 6 — лобковая кость
За грудным отделом позвоночного столба расположен сложный крестец (synsacrum; рис. 106, 8; рис. 107), представляющий собой слившиеся в общую монолитную кость все поясничные (у ворон и голубей 6), все крестцовые (почти у всех видов птиц 2) и часть хвостовых (3—8) позвонков. Сложный крестец неподвижно срастается с последним грудным позвонком. Границы между отделами в сложном крестце провести трудно; крестцовые позвонки (рис. 107, 2) бывают заметны лишь по отверстиям между их поперечными отростками и подвздошными костями. Кости тазового пояса также неподвижно срастаются с позвонками сложного крестца.
Благодаря срастаниям позвонков туловищный участок осевого скелета птиц представляет собой прочное, монолитное образование, обеспечивающее неподвижность туловища, что крайне важно при полете. Образование сложного крестца, слившегося с костями тазового пояса, создает прочную опору для задних конечностей, на которые давит вся тяжесть тела при движении птицы по земле (рис. 107).
Число свободных, подвижно соединенных друг с другом хвостовых позвонков (рис. 106, 9) у птиц невелико (от 5 до 9; у вороны и голубя — обычно 7). Последние хвостовые позвонки (4—8) сливаются в вертикально уплощенную пластинку — копчиковую кость (pygostyle; рис. 106, 10), к боковым поверхностям которой веером прикрепляются основания рулевых перьев. Укорочение хвостового скелета увеличивает общую компактность тела (важно аэродинамически). Характер прикрепления рулевых перьев позволяет использовать хвост в полете (добавочная несущая плоскость) и при посадке (торможение).
Череп. Череп птиц тропибазального типа (мозг не лежит между глазницами). По сравнению с пресмыкающимися резко бросается в глаза значительное увеличение объема мозговой полости, большие глазницы, появление клюва, полное исчезновение зубов, у большинства птиц — заметное смещение вниз большого затылочного отверстия. Кости черепа настолько тонки, что соединение их друг с другом при помощи швов становится невозможным. Поэтому у птиц большинство костей черепа сливается друг с другом; границы между отдельными костями хорошо видны только на черепе птенцов. Тонкие кости и редукция зубов сильно облегчают скелет головы. В то же время благодаря сложной губчатой внутренней структуре кости черепа сохраняют большую прочность.
Череп (cranium) делится на мозговой череп (neurocranium), включающий собственно мозговую (черепную) коробку, носовой отдел и область глазниц, и висцеральный череп (splanchnocranium), к которому относятся челюсти (клюв), кости нёба и подъязычный аппарат.
Рис. 108. Череп вороны
А — сбоку; Б — снизу; В — сверху:
1 — большое затылочное отверстие, 2 — основная затылочная кость, 3 — боковая затылочная кость, 4 — верхняя затылочная кость, 5 — затылочный мыщелок, 6 — теменная кость, 7 —лобная кость, 8 — носовая кость, 9 — чешуйчатая кость, 10 — наружный слуховой проход, 11 — боковая клиновидная кость, 12 — средняя обонятельная кость, 13 — предлобная кость, 14 — предчелюстная кость, 15 — верхнечелюстная кость, 16 — скуловая кость, 17 — квадратно-скуловая кость, 18 — квадратная кость, 19 — основная височная кость, 20 — клювовидный отросток парасфеноида, 21 — сошник, 22 — нёбная кость, 23 — крыловидная кость, 24 — зубная кость, 25 — сочленовная кость, 26 — угловая кость
Начнем рассмотрение черепа с затылочной области. На задне-нижней стенке черепной коробки находится большое затылочное отверстие (foramen occipitale magnum; рис. 108, 1), окруженное четырьмя сросшимися затылочными костями. Вентрально расположена основная затылочная кость (basioccipitale; рис. 108, 2), по бокам — парные боковые затылочные кости (occipitale laterale или exooccipitale; рис. 108, 3). Основная и боковые затылочные кости образуют единственный, как и у пресмыкающихся, затылочный мыщелок (condylus occipitalis; рис. 108,5), сочленяющий череп с первым шейным позвонком. Над затылочным отверстием располагается верхняя затылочная кость (supraoccipitale; рис. 108, 4).
Спереди от верхней и боковых затылочных костей лежат парные теменные кости (parietale; рис. 108, 6). Крышу черепа над глазницами, верхние и задние стенки глазниц и всю переднюю часть мозговой коробки образуют длинные и широкие лобные кости (frontale; рис. 108, 7). Впереди лобных костей у основания клюва лежат парные носовые кости (nasale; рис. 108,  с двумя отростками: верхний идет вперед сбоку от гребня клюва и ограничивает носовое отверстие, сверху, челюстной — вперед и вниз к верхнечелюстной кости и ограничивает носовое отверстие сзади. Боковую часть мозговой коробки образуют парные крупные чешуйчатые кости (squamosum; рис. 108, 5), лежащие книзу от лобных и спереди от теменных и боковых затылочных костей. Верхняя височная дуга у птиц отсутствует. Чешуйчатая кость прикрывает ушные кости, которые срастаются вместе и образуют костные стенки среднего и внутреннего уха; снаружи они обычно не видны. При хорошей препарировке черепа в центре ушного прохода (рис. 108, 10) бывает виден конец тонкой столбчатой косточки (или стремечка, stapes), образовавшийся в результате окостенения подвеска (hyomandibulare) — верхнего элемента подъязычной дуги.
с двумя отростками: верхний идет вперед сбоку от гребня клюва и ограничивает носовое отверстие, сверху, челюстной — вперед и вниз к верхнечелюстной кости и ограничивает носовое отверстие сзади. Боковую часть мозговой коробки образуют парные крупные чешуйчатые кости (squamosum; рис. 108, 5), лежащие книзу от лобных и спереди от теменных и боковых затылочных костей. Верхняя височная дуга у птиц отсутствует. Чешуйчатая кость прикрывает ушные кости, которые срастаются вместе и образуют костные стенки среднего и внутреннего уха; снаружи они обычно не видны. При хорошей препарировке черепа в центре ушного прохода (рис. 108, 10) бывает виден конец тонкой столбчатой косточки (или стремечка, stapes), образовавшийся в результате окостенения подвеска (hyomandibulare) — верхнего элемента подъязычной дуги.
Ниже лобных костей переднюю часть мозговой коробки (являющуюся также задней стенкой глазницы) образуют небольшие парные боковые клиновидные кости (laterosphenoidem; рис. 108, 11). Практически почти всю тонкую межглазничную перегородку образует непарная средняя обонятельная кость (mesethmoideum; рис. 108, 12). Передние стенки глазницы образуют парные (по одной с каждой стороны) предлобные кости (praefrontale; рис. 108, 13); у вороны и других воробьиных птиц они очень малы, и основную часть передней стенки глазницы заполняет отросток средней обонятельной кости.
Клюв птицы состоит из двух частей: надклювья, образованного предчелюстными, верхнечелюстными и носовыми костями, и подклювья. Большую часть надклювья составляют слившиеся парные предчелюстные кости (praemaxillare; рис. 108, 14). Спереди они образуют выпуклую сверху л слегка вогнутую снизу вершину клюва, а назад продолжаются в виде трех парных отростков:
1) лобные отростки соединяются с верхними отростками носовых костей и образуют верхний гребень клюва (ограничивают носовое отверстие сверху и спереди);
2) челюстные отростки отграничивают носовое отверстие сбоку;
3) нёбные отростки образуют костное дно надклювья (они входят в состав твердого нёба).
Небольшие верхнечелюстные кости (maxillare; рис. 108, 15) образуют лишь заднюю нижнюю часть надклювья, соединяясь спереди с челюстными отростками предчелюстных костей; снизу к ним примыкают (сливаясь) челюстные отростки нёбных костей.
От заднего края верхнечелюстной кости отходит назад тонкая костная перекладина, состоящая из двух слившихся костей — скуловой (jugale; рис. 108, 16) и квадратно-скуловой (quadrato-jugale; рис. 108, 17). По своему положению это типичная нижняя дуга, поэтому череп птиц относят к диапсидному типу с редуцированной верхней дугой. Квадратно-скуловая кость причленяется к квадратной кости (quadratum; рис. 108, 18). Нижний конец квадратной кости образует суставную поверхность для причленения нижней челюсти, а другой удлиненный ее конец подвижно сочленяется с чешуйчатой и переднеушной костями.
Посмотрим теперь на череп снизу. В основании черепа, впереди основной затылочной кости (рис. 108, 2) лежит небольшая основная клиновидная кость (basisphenoideum). Она полностью прикрыта широкой основной височной костью (basitemporale; рис. 108, 19) — производной парасфеноида. Передняя часть парасфеноида сохраняется в виде направленного вперед узкого клювовидного отростка (rostrum parasphenoidei; рис. 108, 20). У его переднего конца лежит сошник (vomer; рис. 108, 21). По бокам сошника располагаются хоаны — внутренние отверстия ноздрей.
Нёбные отростки предчелюстных и верхнечелюстных костей сливаются с удлиненными парными нёбными костями (palatinum; рис. 108, 22) и образуют костное дно надклювья. Задние, имеющие сложный профиль, концы нёбных костей налегают на клювовидный отросток парасфеноида. В этом месте к нёбным костям причленяются (суставом) парные крыловидные кости (pterygoideum; рис. 108, 23), задние концы которых также суставом соединяются с квадратными костями.
Рис. 109. Схема кинетизма черепа птицы
Сплошной линией и пунктиром показано изменение положения надклювья при движении костного нёба:
1 — квадратная кость, 2 — крыловидная кость, 3 — нёбная кость, 4 — скуловая кость, 5 — квадратно-скуловая кость, 6 — надклювье, 7 — место перегиба костей
Эти элементы твердого нёба имеют очень важное значение для движения клюва (рис. 109). Если сокращаются мышцы, соединяющие направленный вперед длинный глазничный отросток квадратной кости со стенками глазницы, то направленный вниз конец квадратной кости несколько подвигается вперед и толкает вперед как крыловидные и нёбные кости (место сочленения этих костей может скользить по клювовидному отростку парасфеноида, на который они налегают), так и квадратно- скуловую и скуловую. Давление по этим двум костным мостикам передается на основание надклювья, благодаря чему вершина надклювья приподнимается кверху. При этом прогибаются кости у основания вершины надклювья, в области «переносицы». Перегиб облегчает очень сильное истончение костей в этом месте; у ряда видов здесь сохраняется хрящевая перемычка или даже образуется настоящий сустав. При сокращении мышц, соединяющих череп с нижней челюстью, нижний конец квадратной кости сдвигается назад, оттягивая на себя эти костные связи, и вершина клюва сдвигается книзу.
Описанный сложный костный механизм твердого нёба (основа его — подвижно соединенная с черепом сложной формы квадратная кость) дополняется системой дифференцированных жевательных мышц. Все это обеспечивает возможность довольно разнообразных движений клюва, облегчающих захват добычи, чистку оперения, постройку сложного гнезда и т. п. Возможность дифференцированных движений клюва выработалась, видимо, в связи с преобразованием передних конечностей в крылья, выполняющие только функцию полета.
Подклювье — нижняя челюсть — представляет собой ряд слившихся костей, из которых наиболее крупные зубная (dentale; рис. 108, 24), сочленовная (articulare; рис. 108, 25) и угловая (angulare; рис. 108, 26). Челюстной сустав образуют, соединяясь друг с другом, квадратная и сочленовная кости.
Рис. 110. Подъязычный аппарат вороны:
1 — тело, 2 — его передний отросток, служащий основанием языка, 3 — рожки
Остатки гиоида и жаберных дуг превращаются в подъязычный аппарат (рис. 110). У некоторых птиц (например, дятлов) благодаря большой длине рожков и общей подвижности подъязычного аппарата язык может выдвигаться вперед почти на длину клюва, что облегчает захват добычи.
Конечности и их пояса. Пояс передних конечностей (плечевой пояс) птиц образован парными коракоидами, лопатками и ключицами.
Мощные удлиненные коракоиды (coracoideum; см. рис. 106, 11) своими расширенными нижними концами прочно соединяются малоподвижными суставами с передним краем грудины. Между передними концами коракоидов расположена прикрепляющаяся к ним вилочка (furcula, см. рис. 106, 12), возникшая путем слияния обеих ключиц. Длинные и узкие лопатки (scapula; см. рис. 106, 13) лежат над ребрами; их передние концы прочно соединены со свободными концами коракоидов. В месте соединения лопатки и коракоида расположена довольно глубокая суставная впадина (см. рис. 106, 14), в которую входит головка плечевой кости.
Мощность костей плечевого пояса и их прочное соединение с грудиной обеспечивают опору для крыльев в полете. Удлинение коракоидов увеличивает площадь прикрепления мышц крыла, а также позволяет вынести вперед плечевой сустав, что обеспечивает и удобное складывание крыла в покое, и аэродинамически выгодное его положение в полете, когда центры площадей крыльев лежат на линии, проходящей через центр тяжести птицы. Расположенная между свободными концами коракоидов вилочка обычно играет роль пружины, амортизирующей резкие толчки при взмахах крыла в полете.
Рис. 111. Скелет крыла вороны:
1 — плечевая кость, 2 — лучевая кость, 3 — локтевая кость, 4 —самостоятельные косточки запястья, 5 — пряжка (слившиеся косточки запястья и пясти), 6 — фаланги II пальца, 7 — единственная фаланга I пальца, 8 — единственная фаланга III пальца
Скелет передней конечности, превратившейся в крыло, подвергся значительным изменениям. Плечо (humerus; рис. 111, 1) — мощная трубчатая кость, проксимальный конец которой образует уплощенную головку плеча, входящую в суставную впадину плечевого сустава. Характер суставных поверхностей заметно ограничивает возможность вращательных движений в плечевом суставе, что обеспечивает устойчивость крыла в полете. Мощные эпифизы дистального конца плеча образуют суставные поверхности для сочленения с двумя костями предплечья: более прямой и тонкой лучевой (radius; рис. 111, 2) и более мощной, слегка изогнутой локтевой костью (ulna; рис. 111, 3). На поверхности локтевой кости видны бугорки — места прикрепления очинов второстепенных маховых перьев. Характер суставных поверхностей локтевого сустава обеспечивает прочное соединение костей плеча и предплечья и ограничивает вращательные движения и в этом суставе. При этом сохраняется большая подвижность в одной плоскости — плоскости крыла, что дает возможность птице складывать крыло в покое и изменять его площадь при изменении режима полета.
Особенно резкие изменения в связи с приспособлением к полету произошли в строении кисти. В проксимальном отделе запястья (carpus) сохраняются только две самостоятельные косточки (рис. 111, 4). Они связками почти неподвижно соединяются с костями предплечья. Подвижный сустав расположен между ними и остальными костями запястья, поэтому его называют интеркарпальным. Остальные кости запястья и все кости пясти (metacarpus) сливаются в единое образование— пряжку (carpometacarpus; рис. 111, 5). Резко редуцируется скелет пальцев. Сохраняются две фаланги II пальца (рис. 111, 6), продолжающие ось пряжки. От I пальца сохраняется одна фаланга (рис. 111, 7), к ней прикрепляется пучок перьев, образующий крылышко (alula; см. рис. 98, 4). III палец представлен также одной фалангой (рис. 111, 8), прикрепленной к основанию первой фаланги II пальца.
Эти преобразования в скелете кисти (образование carpometacarpus, редукция пальцев) обеспечивают прочную опору для первостепенных маховых перьев — части крыла, несущей в полете наибольшую нагрузку.
Тазовый пояс состоит из трех пар срастающихся вместе костей: Широкая и длинная подвздошная кость (ilium; см. рис. 106, 15; рис. 107, 4) на большом протяжении срастается со ложным крестцом. К ее наружному краю прирастает седалищная кость (ischium; см. рис. 106, 16; рис. 107, 5), а ниже расположена направленная назад палочкообразная лобковая кость (pubis; см. рис. 106, 17; рис. 107, 6). Все три кости таза образуют вертлужную впадину (acetabulum; см. рис. 106, 18), в которую входит головка бедра.
Большая поверхность тазовых костей и их прочное (неподвижное) соединение с осевым скелетом обеспечивают прочную опору для прикрепления мощных мышц. Эти особенности тазового пояса определяются тем, что передние конечности выполняют только функцию полета, а при движении по земле вся тяжесть тела поддерживается лишь задними конечностями.
Лобковые и седалищные кости правой и левой сторон не соединяются друг с другом на брюшной поверхности (см. рис. 107). Поэтому таз птиц называют открытым; он дает возможность откладывать крупные яйца. Можно предполагать, что открытый таз, увеличивающий подвижность брюшной стенки в тазовой области, способствует интенсификации дыхания в полете.
Рис. 112. Скелет задней конечности вороны:
1 — бедренная кость, 2 — коленная чашечка, 3 — голено-предплюсна (tibio-tarsus), 4 — малая берцовая кость, 5 — цевка (слившиеся кости предплюсны и плюсны), 6 – интертарзальный сустав, 7 — фаланги пальцев; I — IV — пальцы
Скелет задней конечности представлен мощными трубчатыми костями. Бедро (femur; рис. 112, 1) на проксимальном конце имеет хорошо развитую округлую головку для сочленения с тазовым поясом. На дистальном конце образуются рельефные суставные поверхности для сочленения с костями голени. В области коленного сустава в мышечном сухожилии лежит коленная чашечка (patella; рис. 112, 2).
Основной элемент голени — массивная большая берцовая кость, с нижним дистальным концом которой сливаются две проксимальные кости предплюсны; образуется костный комплекс, представляющий собой голено-предплюсну, или тибио-тарзус (tibio-tarsus; рис. 112, 3). Малая берцовая кость (fibula; рис. 112, 4) сильно редуцирована и в виде маленькой тоненькой косточки прирастает к верхней части наружной поверхности большой берцовой кости.
Дистальные элементы предплюсны (tarsus) и все элементы плюсны (metatarsus) сливаются у птиц в единую кость — цевку, или плюсну-предплюсну (tarso-metatarsus; рис. 112, 5). Благодаря этому в конечности появляется добавочный рычаг. Подвижное сочленение располагается между двумя рядами костей предплюсны (между костями приросшими к голени, и костями, слившимися с плюсной), поэтому, как и у пресмыкающихся, этот сустав называется интертарзальным (рис. 112, 6).
На дистальном конце цевки образуются хорошо выраженные суставные поверхности для причленения фаланг пальцев (phalanges digitorum; рис. 112, 7). У подавляющего большинства птиц в задней конечности развиваются четыре пальца, из которых I направлен назад, а II, III, IV — вперед.
Мощные длинные кости скелета задних конечностей, появление добавочного рычага (образование tarso-metatarsus), резкий рельеф суставных поверхностей — все это обеспечивает прочность и подвижность задних конечностей.
ЗАКЛЮЧЕНИЕ
Морфологически класс птиц во многих чертах сохраняет большое сходство со своими предками — пресмыкающимися. На это указывает целый ряд признаков: почти полное отсутствие кожных желез, то, что дистальные отделы задних конечностей покрыты роговыми чешуями, роговой покров клюва, диапсидного типа череп (с редуцированной, однако, верхней дугой), имеющий только один затылочный мыщелок, состав костей черепа, то, что имеются шейные ребра и крючковидные отростки грудных ребер, открытый таз (как у некоторых ископаемых динозавров), большое сходство строения мочеполовой системы и периферических отделов кровеносной системы, присутствие клоаки, размножение откладыванием яиц, сходство эмбрионального развития и т. п.
В процессе эволюции птицы обособились от рептилиеобразных предков путем многих, но часто относительно небольших преобразований морфологических структур, обеспечиваюших существенную интенсификацию физиологических функций и тем самым резко повысивших общий уровень организации птиц по сравнению с пресмыкающимися. Полет и высокая общая подвижность птиц связаны с большим расходом энергии, требующим быстрой компенсации.
Поэтому в эволюции птиц большое значение имело становление особенностей организации, связанных не только с обеспечением физической возможности полета, но и с общим повышением интенсивности обмена веществ.
Возможность полета птиц обеспечивается целым рядом прямых приспособлений. К ним относятся: компактность тела, жесткость и неподвижность туловищного отдела осевого скелета и редукция его хвостового отдела, резкое увеличение размеров грудины и образование на ней киля, прочность пояса передних конечностей (в том числе и образование специального амортизирующего устройства — вилочки), строение скелета крыла (включая и особенности суставных поверхностей и особенно резкие изменения в строении кисти), мощность мускулатуры, обеспечивающая движения крыла, развитие перьев — легких и прочных роговых образований, создающих несущие плоскости крыльев и хвоста и обеспечивающих обтекаемость тела в по лете, редукция зубов, облегчающая скелет черепа, общее увеличение объема головного мозга и особенно сильное развитие мозжечка — центра равновесия и координации движений, увеличение массы нервных клеток спинного мозга в области крыльев и т. д.
Клюв с его подвижным скелетным механизмом и дифференцированной мускулатурой стал очень тонко действующим инструментом, способным к сложным движениям. Образование клюва и одновременно увеличение длины и подвижности шеи освободило передние конечности от многих добавочных функций (захват и удерживание добычи, чистка покровов и т. п.) и тем самым способствовало их превращению только в орган полета — крылья. Удлинение шеи и возрастание ее подвижности компенсировало неподвижность туловища и дало возможность, изменяя положение головы, в каких-то пределах менять положение центра тяжести тела, что важно при изменении режима полета.
Преобразование передних конечностей в орган полета — крылья сопровождалось укреплением тазового пояса и скелета задних конечностей, а также возрастанием массы их мышц, обеспечивающих возможность «двуногого» движения на земле, по ветвям деревьев и т. п.
Полет требует затраты большого количества энергии. Например у голубя, летящего со скоростью 70 км/ч, расход энергии в 27 раз больше, чем в покое. Резкое возрастание энергетических возможностей обеспечивается изменениями многих систем органов.
Легкие птиц относительно малы по объему, но велика их губчатость (и тем самым большая поверхность окисления). Сложная система воздушных мешков, связанная с легкими, занимает до 20% объема тела — она резко увеличивает объем вдыхаемого воздуха и дает возможность насыщать кровь кислородом и во время вдоха, и во время выдоха. Чем интенсивнее полет, тем чаще дыхание1, что обеспечивает более интенсивное насыщение крови кислородом и удаление из нее углекислоты при ускорении движения. Одновременно воздушные мешки осуществляют интенсивную теплоотдачу во время полета, предохраняя организм летящей птицы от перегрева.
Полное разделение большого и малого кругов кровообращения резко увеличивает возможность насыщения тканей кислородом. Сердце у птиц относительно очень велико — составляет 0,8—2,5% от веса тела (у рептилий 0,2—0,3%; у млекопитающих 0,2—1,5%). Относительно очень велика у птиц и частота сердцебиений: сердце крупных птиц сокращается 140—200 раз в 1 мин, мелких — до 500—600. В полете частота сердцебиений резко возрастает: так, у голубя в покое около 170 сердцебиений в 1 мин, а в полете 350—600; у мелких птиц частота пульса в полете до 1000 и более. Большой объем сердца и высокая частота сердцебиений создают быструю циркуляцию крови по организму, обеспечивая интенсивное насыщение тканей и органов питательными веществами и кислородом и освобождение их от углекислоты и продуктов распада.
Дифференцировка желудка на два отдела — железистый и мускульный — обеспечивает механическую переработку пищи (в мускульном желудке может развиваться давление до 20—30 кг на 1 см2), что значительно увеличивает интенсивность ее переваривания. Этому же помогает свойственная ряду видов птиц способность отбрасывания погадок, т. е. удаления непереваренных остатков пищи (хитин, шерсть, кости и т. п.) из желудка через пищевод.
Обеспечивает интенсивное пищеварение и высокая активность пищеварительных ферментов. Поэтому скорость переваривания пищи у птиц заметно выше, чем у пресмыкающихся и млекопитающих. Так, воробей переваривает гусеницу за 15 мин, жука — за 1 ч, зерно — за 3—4 ч (считая от времени принятия пищи до вывода ее непереваренных остатков). Интенсивное пищеварение позволяет перерабатывать большое количество пищи, обеспечивая высокие энергетические потребности организма. Особенности пищеварительной системы и органов захвата пищи наряду с большой подвижностью, высоким уровнем высшей нервной деятельности и совершенством органов чувств способствовали образованию широких пищевых связей и создали возможность использования очень большого числа кормовых объектов.
Интенсивность пищеварения, кровообращения и дыхания обеспечивает высокий уровень обмена веществ, что наряду с образованием высоко эффективного теплоизолирующего покрова из пуха и перьев приводит к теплокровности птиц (гомойотермия), т. е. к их способности поддерживать температуру тела на постоянном уровне независимо от изменения температуры окружающей среды. Косвенным показателем высокого уровня обмена веществ птиц может служить температура их тела, которая у крупных видов равна 38—40°С, а у мелких видов достигает 41—43°С.
Значительное увеличение объема головного мозга и возрастание числа нервных клеток в нем, по сравнению с рептилиями, определяют — более высокий уровень высшей нервной деятельности птиц. Это проявляется и в очень сложных системах врожденных рефлексов, определяющих поведение каждой особи, и в значительном возрастании роли индивидуального опыта, т. е. в выработке условных рефлексов.
Плодовитость птиц меньше, чем у пресмыкающихся. Увеличение размеров яиц (откладка которых обеспечивается открытым тазом) увеличивает размеры вылупившегося детеныша. Сопутствующие размножению сложные биологические явления (постройка гнезда, насиживание, выкармливание, обогрев и охрана птенцов и т. п.) обеспечивают ускорение эмбрионального и постэмбрионального развития и повышенное выживание потомства.
Совершенство органов движения (большая подвижность), гомойотермия, сложность высшей нервной деятельности и совершенство органов чувств (особенно зрения и слуха) позволили птицам широко расселиться по всему земному шару. Практически нет на земле ни одного района, включая высокие широты Арктики и Антарктики, где не встречались бы те или иные виды птиц.
Более высокий уровень организации птиц, по сравнению с пресмыкающимися, находит свое отчетливое выражение не только в значительно более широком их распространении по всему земному шару, но и в более сложных и разнообразных взаимоотношениях со средой, с условиями жизни. Так, при сезонных неблагоприятных изменениях условий жизни (наступление зимы) пресмыкающиеся и земноводные прекращают свою активность, впадают в состояние оцепенения. У птиц в целом реакции при наступлении неблагоприятных сезонов года многообразнее и шире. Часть птиц совершает миграции (протяженностью до 5—10 тыс. км и более), улетая на зимовки в благоприятные по погодным и кормовым условиям районы. Однако значительное число видов остается в умеренных и северных широтах в течение всего года. Переживать суровые условия зимы им помогают довольно разнообразные приспособления.
Для многих видов свойственна резко выраженная сезонная смена кормов: переход зимой на питание менее калорийными, но массовыми кормами (например, тетерев летом питается насекомыми, ягодами, зелеными частями растений, зимой — преимущественно почками и сережками некоторых пород деревьев). Неперелетные (оседлые) птицы совершают небольшие перемещения, концентрируясь в участках с большим количеством пищи (глухарь в сосновых лесах, где питается хвоей сосны; синицы, вороны — в населенных пунктах и т. п.). Некоторые виды осенью запасают пищу, пряча ее в укромных местах (кедровки, сойки, поползни — орехи лещины, «орешки» кедра, желуди; синицы — семена деревьев и кустарников). Оседлые птицы во время осенней линьки получают перьевой покров с лучшими теплоизолирующими свойствами, чем летний (большее развитие пуховой части пера, большее число перьев и пуха на единицу поверхности тела и т. п.); у некоторых видов зимний наряд резко отличается по окраске, помогая маскировке (белая куропатка).
У части видов отчетливо выражены и приспособительные особенности поведения: ночевки в укрытиях (тетеревинные и некоторые другие птицы — в снегу; мелкие воробьиные — в дуплах, щелях за корой и т. п.). При этом птицы некоторых видов в сильные морозы ночуют группами, взаимно обогревая в убежище друг друга (синицы, воробьи и др.).
Несмотря на резкое повышение уровня жизнедеятельности и широкие экологические возможности, открываемые гомойотермией, отчетливая специализация птиц к полету в известной мере ограничивает круг экологических приспособлений этих животных. Отсутствие столь узкой специализации у млекопитающих создало предпосылки к возникновению в процессе эволюции этого класса значительно большего разнообразия жизненных форм и более многообразных взаимоотношений со средой.
Дополнительная литература
Гладков Н. А., Дементьев Г. П., Птушенко Е. С., Судиловская А. М. Определитель птиц СССР. М., 1964.
Гуртовой Н. Н., Матвеев Б. С, Дзержинский Ф. Я. Практическая зоотомия позвоночных. Птицы, млекопитающие. М., 1980.
Дементьев Г. П. Птицы. Руководство по зоологии, т. 6. М., 1940.
Дементьев Г. П. Птицы нашей страны. М., 1962.
Иванов А. И., Штегман Б. К. Краткий определитель птиц СССР. Л., 1964.
Карташев Н. Н. Систематика птиц. М., 1974.
Михеев А. В. Биология птиц. М., 1960.
Шмальгаузен И. И. Основы сравнительной анатомии. М., 1947.
Шульпин Л. М. Орнитология. Л., 1940.
НазадОглавлениеДалее
Руководство по зоологии [Текст] / Сост. Г. Г. Абрикосов, В. Н. Беклемишев, В. А. Догель, Л. А. Зенкевич, Н. А. Ливанов, С. Д. Муравейский, Е. Н. Павловский, П. Д. Резвой ; Под ред. Л. А. Зенкевича ; Науч.-исслед. ин-т зоологии Моск. гос. ун-та
Карточка
Руководство по зоологии [Текст] / Сост. Г. Г. Абрикосов, В. Н. Беклемишев, В. А. Догель, Л. А. Зенкевич, Н. А. Ливанов, С. Д. Муравейский, Е. Н. Павловский, П. Д. Резвой ; Под ред. Л. А. Зенкевича ; Науч.-исслед. ин-т зоологии Моск. гос. ун-та. — Москва ; Ленинград : Гос. изд-во биол. и мед. литературы, 1937-1951. — 8 т.; 27 см.
Т. 6: Позвоночные. Птицы. Т. 6 : Сост. Дементьев / Сост. Дементьев. — 1940. — 856 с., 1 л. ил. : ил.
Т. 6: Позвоночные. Птицы. Т. 6 : Сост. Дементьев / Сост. Дементьев. — 1940. — 856 с., 1 л. ил. : ил.
Шифр хранения:
FB ОПБ
FB И/Ру-489
FB R 386/80
FB R 54/273
FB R 54/278
OMF Ф 1-72/13553
FB R 388/67
Описание
| Заглавие | Руководство по зоологии [Текст] / Сост. Г. Г. Абрикосов, В. Н. Беклемишев, В. А. Догель, Л. А. Зенкевич, Н. А. Ливанов, С. Д. Муравейский, Е. Н. Павловский, П. Д. Резвой ; Под ред. Л. А. Зенкевича ; Науч.-исслед. ин-т зоологии Моск. гос. ун-та |
|---|---|
| Коллекции ЕЭК РГБ | Каталог документов с 1831 по настоящее время |
| Дата поступления в ЭК | 10.02.2016 |
| Каталоги | Книги (изданные с 1831 г. по настоящее время) |
| Выходные данные | Москва ; Ленинград : Гос. изд-во биол. и мед. литературы, 1937-1951 |
| Физическое описание | 8 т.; 27 см |
| Том | Т. 6: Позвоночные. Птицы. Т. 6 : Сост. Дементьев / Сост. Дементьев. — 1940. — 856 с., 1 л. ил. : ил. |
| Язык | Русский |
| Места хранения | FB ОПБ |
| FB И/Ру-489 | |
| FB R 386/80 | |
| FB R 54/273 | |
| FB R 54/278 | |
| OMF Ф 1-72/13553 | |
| FB R 388/67 |